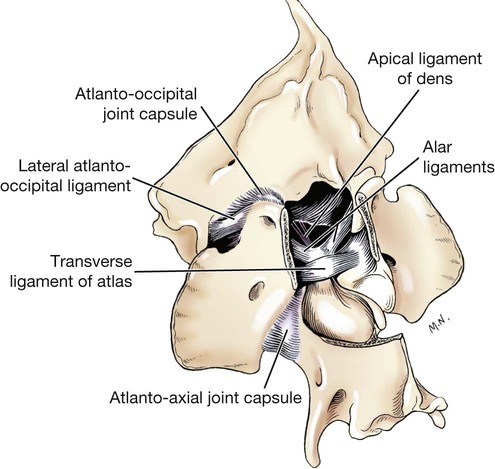
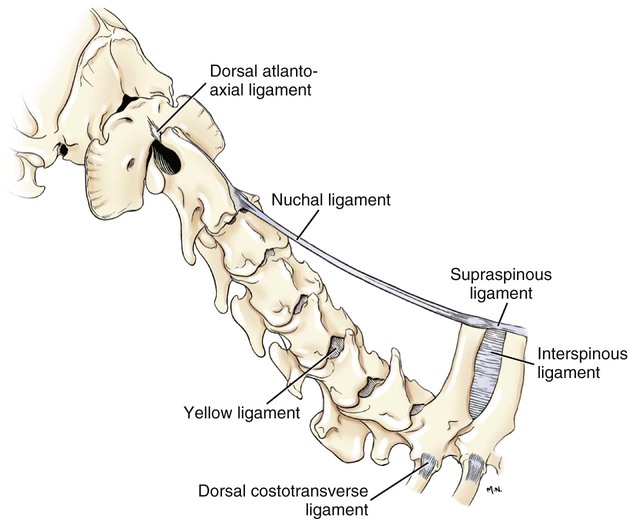
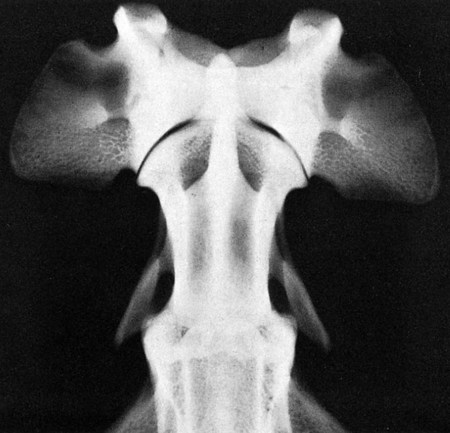
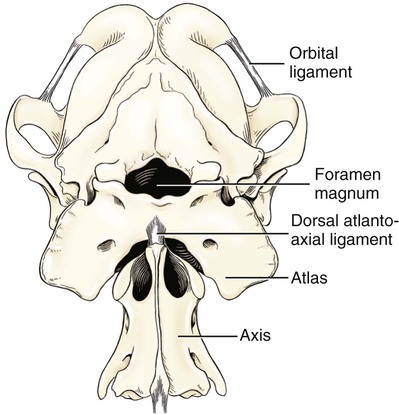
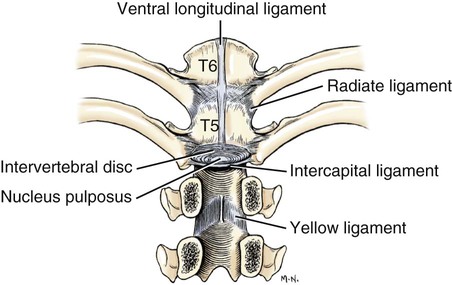
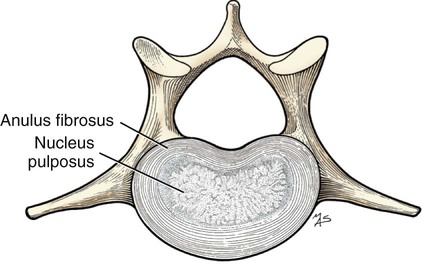
Articulations, or joints (articulationes [juncturae] ossium), are formed when two or more bones are united by fibrous, elastic, or cartilaginous tissue or by a combination of these tissues. Three main groups are recognized and named according to their most characteristic structural features. Where little movement is required, the union is short, direct, and often transitory. A fibrous joint (junctura fibrosa), formerly known as a synarthrosis, is one of this nature. Such joints include syndesmoses, sutures, and gomphoses. A cartilaginous joint (junctura cartilaginea), formerly known as an amphiarthrosis, permits only limited movement, such as compression or stretching. A synovial joint (junctura synovialis) formerly known as a diarthrosis or true joint, facilitates mobility. The studies of Kadletz (1932) provide detailed information on the arthrology of the dog, and the well-documented work of Barnett et al. (1961) discusses the structure and mechanics of synovial joints in considerable detail. At the Tokyo meeting of the International Nomenclature Committee, arthrologia was adapted as the most appropriate heading and articulatio replaced junctura. The sixth edition of Nomina Anatomica (1989), retained arthrologia and articulatio. It should be noted that the discarded original term, syndesmologia, for all joints is similar sounding to the term syndesmosis, which is used to denote one type of fibrous joint. The nerve supply of synovial joints is derived from cutaneous or muscular branches in the vicinity of the joint. Included in these articular nerves are proprioceptive fibers, nociceptor fibers and sympathetic visceral efferent and visceral afferent fibers related to vasomotor or vasosensory functions respectively. Some areas of the joint capsule are more richly innervated than others. Four types of joint receptors are present in most animal joints (Polacek, 1966; Zimny, 1988): (1) Ruffini-like receptors in the capsule, (2) Pacinian-like receptors in the capsule, (3) Golgi tendon organs in ligaments, and (4) free nerve endings. If a joint has intraarticular structures, they are usually innervated. The purpose of the innervation is proprioception and the recognition of angular movement; thus posture is very dependent on these endings. It is likely that the stifle joint with its many ligaments and menisci has the richest innervation of all joints. O’Connor and Mc-Connaughey (1978) and O’Connor (1976, 1984) found both Ruffini endings and Pacinian corpuscles in the menisci of the dog and cat. Sfameni (1902) found single or grouped nerve endings that arose from single axons in the dog and suggested the name Ruffini endings because they resembled those described in the skin by Ruffini in 1894. Gardner (1950) reviewed the morphologic and physiologic characteristics of joints in the human, including their innervation, and cites more than 500 references. Ansulayotin (1960) studied the nerves that supply the appendicular joints in the dog. The synovial fluid (synovia) serves chiefly to lubricate the contact surfaces of synovial joints. In all cases these surfaces are hyaline cartilage or fibrocartilage. Fibrocartilage contains few blood vessels and nerves, and hyaline cartilage has neither. Therefore the synovial fluid serves the additional function of transporting nutrient material to the hyaline cartilage and removing the waste metabolites from it. Synovia also enables the wandering leukocytes to circulate in the joint cavity and phagocytize the products of the wear and tear of the articular cartilage. In many joints there is little, if any, free synovia. The average volume in the stifle joint of adult dogs of various sizes varies from 0.2 mL to 2 mL. The general health and condition of the dog has a marked influence on the amount of synovia present in the joints. Synovia is thought to be a dialysate, although mucin is probably produced by the fibroblasts of the synovial membrane (Davies, 1944). The chemical composition of synovia closely resembles that of tissue fluid. In addition to mucin, it contains salts, albumin, fat droplets, and cellular debris. The quantitative composition of synovia depends largely on the type of tissue underlying the surface fibroblasts and the degree of vascularity of this tissue. Because of its mucin content, the synovia forms a viscous capillary film on the articular cartilage. The articular cartilage (cartilago articularis) is usually hyaline cartilage. It covers the articular surfaces of bones where its deepest part may be calcified. It contains no nerves or blood vessels, although it is capable of some regeneration after injury or partial removal (Bennett et al., 1932). It receives its nutrition from the synovia. The articular cartilage varies in thickness in different joints and in different parts of the same joint. It is thickest in young, healthy joints and in joints that bear considerable weight. Its thickness in any particular joint is in direct proportion to the weight borne by the joint, and it may atrophy from disuse. Healthy articular cartilage is translucent, with a bluish sheen. Elasticity and compressibility are necessary physical properties that it possesses. This resiliency guards against fracture of bone by absorbing shock. A meniscus (meniscus articularis), or disc (discus articularis), is a complete or partial fibrocartilaginous plate that divides a joint cavity into two parts. The temporomandibular joint contains a thin, but complete, articular disc, and, because the capsular ligament attaches to the entire periphery of the disc, the joint cavity is completely divided into two parts. Two menisci are found in the stifle joint, and neither is complete, thus allowing all parts of the joint cavity to intercommunicate. Menisci have a small blood and nerve supply and are capable of regeneration (Dieterich, 1931). Their principal function, according to MacConaill (1932), is “to bring about the formation of wedge-shaped films of synovia in relation to the weight-transmitting parts of joints in movement.” An obvious function is the prevention of injury from concussion. The stifle and temporomandibular joints are the only synovial joints in the dog that possess menisci, or discs. Synovial joints may be classified according to (1) the number of articulating surfaces involved, (2) the shape or form of the articular surfaces, or (3) the function of the joint (Barnett et al., 1961). The classification of synovial joints (Nomina Anatomica Veterinaria, 2005) is based on the shape or form of the articular surfaces. There are seven basic types: The temporomandibular joint (articulatio temporomandibularis) (Figs. 5-1 and 5-2) is a condylar joint that allows considerable sliding movement. The transversely elongated condyle of the mandible does not correspond entirely to the articular surface of the mandibular fossa of the temporal bone. A thin articular disc (discus articularis) lies between the cartilage-covered articular surface of the condyloid process of the mandible and the similarly covered mandibular fossa of the temporal bone. The loose joint capsule extends from the articular cartilage of one bone to that of the other. On the temporal bone the capsular ligament also attaches to the retroarticular process. It attaches to the entire edge of the disc as it passes between the two bones. The joint cavity is thus completely divided into a dorsal compartment, between the disc and temporal bone, and a ventral compartment, between the disc and mandible. Laterally the fibrous part of the joint capsule is strengthened by fibrous strands to form the lateral ligament (lig. laterale). The lateral ligament becomes progressively tighter as the jaws open, and if one or the other is unduly lax owing to stretching or joint dysplasia, it is possible to dislocate the temporomandibular joint. Robins and Grandage (1977) described open-mouth jaw locking and its surgical correction in two Basset Hounds with temporomandibular joint dysplasia. Differential movement at the joints, when the jaws were opened widely, allowed locking of the coronoid process lateral to the zygomatic arch. Vollmerhaus and Roos (1996) described transverse movement of the temporomandibular joint in 20 dogs of various breeds. This movement is important for mastication. Umphlet et al (1988) described the effect of hemimandibuloectomy on the joint. Scapino (1965, 1981) investigated the morphologic characteristics and function of the intermandibular articulation in the dog and other carnivores. He described four types of articulations, ranging from flexible to synostosed. He considers the dog to have a flexible joint that permits a moderate amount of independent movement of the mandibles and found this to be the most common type of union in carnivores. When the mandibles of such a joint are separated, the articular surfaces are flat or have low rugosities. A smooth area can be seen rostrodorsally, and the articular space is usually wider caudally than rostrally. The joint is characterized by a single fibrocartilage pad, cruciate ligaments, and a venous plexus. In the wolf and dog the articulation is not stiff, as in the lion and tiger, and is not synostosed, as in the badger and panda. The joints of the auditory ossicles (articulationes ossiculorum auditus) allow for movement of the malleus, incus, and stapes (see Chapter 20). The head of the malleus articulates with the body of the incus via a synovial incudomallear joint (articulatio incudomallearis). The lenticular process of the long crus of the incus, with the head of the stapes, likewise forms a synovial joint, which is called the incudostapedial joint (articulatio incudostapedia). The footplate, or base, of the stapes attaches to the margin of the vestibular window (fenestra vestibuli) by means of a fibrous union (syndesmosis tympanostapedia). There is a common joint cavity formed by the articulations of the occipital condyles with the atlas, and the atlas with the axis. By means of silicone casts this cavity (Fig. 5-3) has been studied and described as the “composite occipito-atlas-axis joint cavity” by Watson et al. (1986). A cast of the cavity resembles the shape of an “hourglass” with the ends removed or “popeye” holding up the head. It appears to be a composite of five synovial joints: right and left atlantooccipital joints, a median joint cavity between the ventral articular surface of the dens and the dorsal surface of the ventral arch of the atlas, and right and left atlantoaxial joints. The synovial bursa between the transverse atlantal ligament and the dens does not communicate with the common joint cavity. The lateral ligament (lig. laterale) of the atlantooccipital joint (see Fig. 5-5) runs from the lateral part of the dorsal arch of the atlas to the paracondylar process of the occipital bone. Its course is cranioventrolateral, and its caudal attachment is narrower than its cranial one. Another small ligament runs from each side of the inner surface of the lateral part of the ventral arch of the atlas to the lateral part of the foramen magnum. Ventral and medial to these ligaments the unpaired joint cavities between the skull and the atlas and between the atlas and the axis freely communicate. The atlantoaxial joint (articulatio atlantoaxialis) (see Figs. 5-3 to 5-6) is a pivot joint that permits the head and atlas to rotate around a longitudinal axis. The joint capsule is loose and uniformly thin as it extends from the dorsal part of the cranial articular surface of one side of the axis to a like place on the opposite side. Cranially it attaches to the caudal margins of the caudal articular foveae and ventral arch of the atlas. The fibrous layer of the joint capsule extends from right to left between the dorsal arch of the atlas and the arch of the axis. This is the dorsal atlantoaxial membrane, or membrana tectoria. The apical ligament of the dens (lig. apicis dentis) (see Fig. 5-5) leaves the apex of the dens and passes straight cranially to the basioccipital bone at the ventral part of the foramen magnum. The apical ligament represents a remnant of the notochord. The two alar ligaments (ligg. alaria) are wider and heavier than the apical ligament. They attach to the dens on either side of the apical ligament and diverge from each other to attach to the occipital bone medial to the caudal parts of the occipital condyles. The transverse atlantal ligament (lig. transversum atlantis) is a thick ligament that connects one side of the ventral arch of the atlas to the other. It crosses dorsal to the dens and functions to hold this process against the ventral arch of the atlas. A spacious bursa exists between the ventral surface of the ligament and the dens. Atlantoaxial subluxation with absence of the dens has been reported frequently, particularly in toy breeds, and has been ascribed to either congenital developmental or degenerative causes. Injury may result in a fracture of the dens. In almost all instances there is a tilting or dorsal displacement of the axis into the vertebral canal, with resultant compression of the spinal cord (Cook & Oliver, 1981; Oliver & Lewis, 1973). The nuchal ligament (lig. nuchae) (Fig. 5-7) is composed of longitudinal yellow elastic fibers that attach cranially to the caudal part of the large spinous process of the axis. It extends caudally to the dorsal extremity of the spinous process of the first thoracic vertebra. It is a laterally compressed, paired band that lies between the medial surfaces of the mm. semispinales capiti. The yellow nature of the nuchal ligament continues caudally in the supraspinous ligament to the tenth thoracic spinous process (Baum & Zietzschmann, 1936). The supraspinous ligament (lig. supraspinale) (see Figs. 5-12 and 5-7) extends from the spinous process of the first thoracic vertebra caudally to the third caudal vertebra. It is a thick band especially in the thoracic region, where it attaches to the apices of the spines as it passes from one to another. Bilaterally the dense collagenous thoracolumbar fascia imperceptibly blends with it throughout the thoracic and lumbar regions. The thin interspinous ligaments send some strands to its ventral surface, but the supraspinous ligament more than the interspinous ligaments prevents abnormal separation of the spines during flexion of the vertebral column (Heylings 1980). The ventral longitudinal ligament (lig. longitudinale ventrale) (Fig. 5-8) lies on the ventral surfaces of the bodies of the vertebrae. It can be traced from the axis to the sacrum, but it is best developed caudal to the middle of the thorax. The dorsal longitudinal ligament (lig. longitudinale dorsale) (see Fig. 5-10) lies on the dorsal surfaces of the bodies of the vertebrae. It therefore forms a part of the floor of the vertebral canal. It is narrowest at the middle of the vertebral bodies and widest over the intervertebral fibrocartilages. The dorsal longitudinal ligament attaches to the rough ridges on the dorsum of the vertebral bodies and to the intervertebral fibrocartilages. It extends from the dens of the axis to the end of the vertebral canal in the caudal region. The dorsal longitudinal ligament is thicker than the ventral longitudinal ligament. The intervertebral discs (disci intervertebrales) are interposed in every intervertebral space (except between C1 and C2), uniting the bodies of the adjacent vertebrae (Figs. 5-8 and 5-9). In the sacrum of young specimens, transverse lines indicate the planes of fusion of the discs with the adjoining vertebral bodies. The thickness of the discs is greatest in the cervical and lumbar regions, the thickest ones being between the last few cervical vertebrae. The thinnest discs are in the caudal region. Those between the last few segments being smaller in every way than any of the others. Each intervertebral disc consists of an outer laminated fibrous ring, the anulus fibrosus and a central, amorphous, gelatinous center, the nucleus pulposus. The nucleus pulposus of a young dog is proportionally larger than that of an adult and more mucoid than fibroid for 1 to 7 years (King & Smith, 1955). It is a mass of mesodermal cell remnants of the notochord in a homogeneous basophilic intercellular material. Eventually small foci of degeneration and fibrosis occur, which make the disc appear opaque rather than gelatinous and may obscure the boundary with the annulus fibrosus. In chondrodystrophic breeds a chondroid degeneration may occur in young adults that eventually calcifies. There may be ossification within the disc without any adverse effect on surrounding tissues. However, the loss of function of the nucleus pulposus may result in tearing of the anulus fibrosis dorsally with protrusion or extrusion of degenerate nuclear material into the vertebral canal. This is a common cause of discomfort with or without neurologic deficits caused by spinal cord compression in these chondrodystrophic breeds. A similar but fibroid degeneration may occur in nonchondrodystrophic breeds at an older age.
Arthrology
General
Synovial Joints
Structure of Synovial Joints
Classification of Synovial Joints
Ligaments and Joints of the Skull
Temporomandibular Joint
Intermandibular Joint
Joints of Auditory Ossicles
Ligaments and Joints of the Vertebral Column
Atlantooccipital Articulation
Atlantoaxial Articulation
Long Ligaments of the Vertebral Column
Intervertebral Discs and Short Ligaments of the Vertebral Column
< div class='tao-gold-member'>
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Arthrology

Only gold members can continue reading. Log In or Register to continue
