Chapter 12 Vestibular System
Special Proprioception
The vestibular system is the primary sensory system that maintains the animal’s balance, its normal orientation relative to the gravitational field of the earth. This orientation is maintained in the setting of linear or rotatory acceleration or deceleration or tilting of the animal. The vestibular system is responsible for maintaining the position of the eyes, neck, trunk, and limbs relative to the position or movement of the head at any time.
ANATOMY AND PHYSIOLOGY
Receptor
These membranous and bony labyrinths are formed adjacent to the first and second branchial arches and their corresponding first pharyngeal pouch and first branchial groove. The first branchial groove gives rise to the external ear canal. The first pharyngeal pouch forms the auditory tube and the mucosa of the middle-ear cavity. The intervening tissue forms the tympanum. The ear ossicles are derived from the neural crest of branchial arches 1 (malleus and incus) and 2 (stapes). These ossicles become components of the middle ear associated laterally with the tympanum (malleus) and medially with the vestibular window of the bony labyrinth of the inner ear (stapes).
Anatomically, the bony labyrinth in the petrous part of the temporal bone consists of three continuous fluid-filled portions (Figs. 12-1 and 12-2). These areas are the large vestibule and the three semicircular canals and the cochlea, which arise from the vestibule. Dilation in one end of each of the bony semicircular canals is the ampulla. All three continuous bony components contain perilymph, a fluid similar to cerebrospinal fluid (CSF), from which it may be derived. In the bony labyrinth are two openings: the vestibular and cochlear windows, which are named according to the components of the bony labyrinth in which they are located. Each opening is covered by a membrane, and the stapes is inserted in the membrane that covers the vestibular window.
The ectodermally derived membranous labyrinth consists of four fluid-filled compartments, all of which communicate (Fig. 12-3 see also Figs. 12-1 and 12-2). These compartments are contained within the components of the bony labyrinth and include the saccule and utriculus within the bony vestibule, the three semicircular ducts within the bony semicircular canals, and a cochlear duct within the bony cochlea. The endolymph contained within the membranous labyrinth is thought to be derived from the blood vessels along one wall of the cochlear duct and is absorbed back into the blood through the blood vessels surrounding the endolymphatic sac. The three semicircular ducts are the anterior (vertical), posterior (vertical), and lateral (horizontal). Each semicircular duct is oriented at right angles to the others. Thus rotation of the head around any plane causes endolymph to flow within one or more of the ducts. Each semicircular duct connects at both ends with the utriculus, which, in turn, connects with the saccule by way of the intervening endolymphatic duct and sac. The saccule connects with the cochlea duct by the small ductus reuniens.
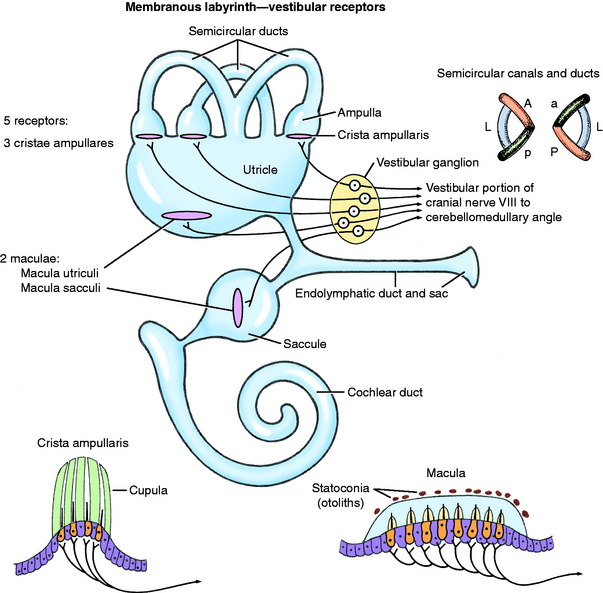
Crista Ampullaris
At one end of each membranous semicircular duct is a dilation called the ampulla. On one side of the membranous ampulla, a proliferation of connective tissue forms a transverse ridge called the crista (see Figs. 12-1 through 12-3). It is lined on its internal surface by columnar neuroepithelial cells. On the surface of the crista is a gelatinous structure that is composed of a protein-polysaccharide material called the cupula, which extends across the lumen of the ampulla. This neuroepithelium is composed of two basic cell types: hair cells and supporting cells. The neurons of the vestibulocochlear nerve are derived from otic placode ectoderm. The dendritic zones of the neurons of the vestibular portion of the vestibulocochlear nerve are in synaptic contact with the base of the hair cells. These hair cells have on their luminal surface 40 to 80 hairs, or modified microvilli (stereocilia), and a single modified cilium (kinocilium). These structures project into the overlying cupula. Movement of fluid in the semicircular ducts causes deflection of the cupula, which is oriented transversely to the direction of flow of the endolymph. This deflection bends the stereocilia, which is the source of the stimulus by way of the hair cells to the dendritic zone of the vestibular neuron that is in synaptic relationship with the plasmalemma of the hair cell.
The vestibular neurons are tonically active, and their activity is excited or inhibited by deflection of the cupula in different directions. Each semicircular duct on one side is paired with a semicircular duct on the opposite side by their common position in a parallel plane. These synergistic pairs are the left and right lateral ducts, the left anterior and right posterior ducts, and the left posterior and right anterior ducts. When movement in the direction of one of these three planes stimulates the vestibular neurons of the crista of one duct, they are inhibited in the opposite duct of the synergistic pair. For example, rotation of the head to the right causes the endolymph to flow in the right lateral duct such that the cupula is deflected toward the utriculus and the cupula of the left lateral duct is deflected away from the utriculus. This action causes increased activity of vestibular neurons on the right side and decreased activity on the left side, resulting in a jerk nystagmus to the right side, which is an involuntary rhythmic oscillation of the eyes. The anatomic orientation of the stereocilia relative to the kinocilium on the surface of the crista is responsible for the difference in activity relative to the direction of the cupula deflection. Deviation of the stereocilia toward the kinocilium increases vestibular neuron activity. These receptors are not affected by a constant velocity of movement but respond to acceleration or deceleration, especially when the head is rotated.
Macula
The macula is the receptor found in the utriculus and saccule, which are located in the bony vestibule. These maculae are on one surface of each of these saclike structures (see Figs. 12-1 through 12-3). Each macula is an oval-shaped plaque in which the membranous labyrinth has proliferated. The surface of the macula consists of columnar epithelial cells. This neuroepithelium is composed of hair cells and supporting cells. Covering the neuroepithelium is a gelatinous material, the statoconiorum (otolithic) membrane. On the surface of this membrane are calcareous crystalline bodies known as statoconia (otoliths). Similar to the hair cells of the cristae, the macular hair cells have projections of their luminal cell membranes—stereocilia and kinocilia—into the overlying statoconiorum membrane. Movement of the statoconia away from these cells is the initiating factor in bending the stereocilia to stimulate an impulse in the dendritic zones of the vestibular neurons that are in synaptic relationship with the base of the hair cells. The macula in the saccule is oriented in a vertical direction (sagittal plane), whereas the macula of the utriculus is in a horizontal direction (dorsal plane). Thus gravitational forces continually affect the position of the statoconia relative to the hair cells. These structures are responsible for the sensation of the static position of the head and linear acceleration or deceleration. They function in static equilibrium. The macula of the utriculus may be more important as a receptor for sensing changes in head posture, whereas the macula of the saccule may be more sensitive to vibrational stimuli and loud sounds.
Vestibulocochlear Nerve: Cranial Nerve VIII—Vestibular Division
The dendritic zone of the vestibular portion of cranial nerve VIII is in a synaptic relationship with the hair cells of each crista ampullaris and the macula utriculi and macula sacculi. The axons course through the internal acoustic meatus with those of the cochlear division of this nerve. The cell bodies of these bipolar-type sensory neurons are inserted along the course of the axons within the petrous portion of the temporal bone, where they form the vestibular ganglion (see Fig. 12-3). After leaving the internal acoustic meatus with the cochlear division of the vestibulocochlear nerve, the vestibular nerve axons pass to the lateral surface of the rostral medulla at the cerebellomedullary angle, which occurs at the level of the trapezoid body and the attachment of the caudal cerebellar peduncle to the cerebellum. The vestibular nerve axons enter the medulla between the caudal cerebellar peduncle and the spinal tract of the trigeminal nerve and terminate in telodendria at one of two sites. The majority of them terminate in the vestibular nuclei in the medulla and pons. A few course directly into the cerebellum by way of the caudal peduncle and terminate in the fastigial nucleus in the cerebellar medulla and the cortex of the flocculonodular lobe. These latter axons form the direct vestibulocerebellar tract.
Vestibular Nuclei
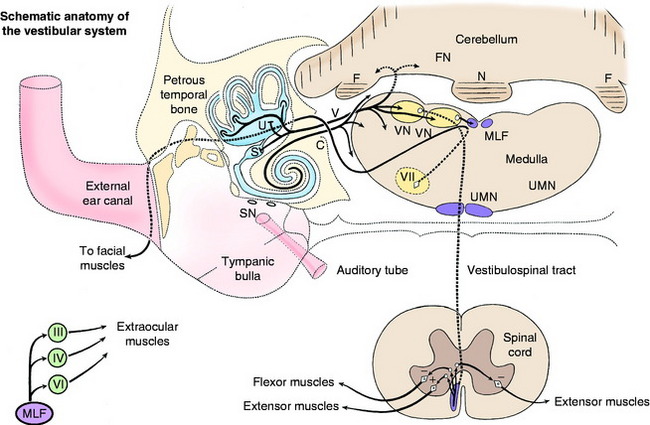
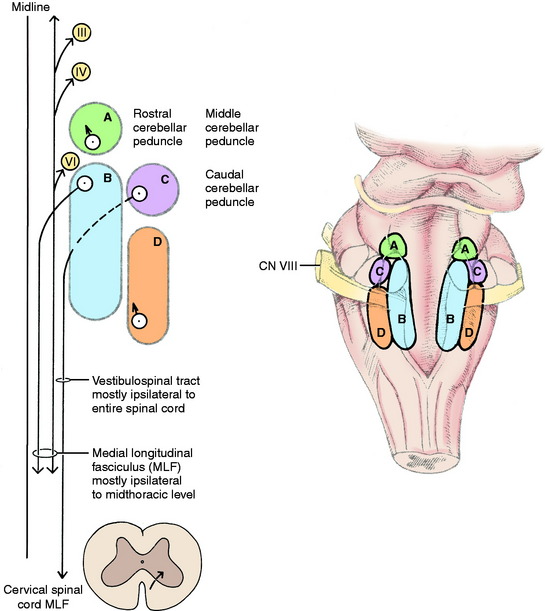
On either side of the dorsal part of the pons and medulla adjacent to the lateral wall of the fourth ventricle are four vestibular nuclei (Fig. 12-4 see also Fig. 12-2). From the level of the rostral and middle cerebellar peduncles, they extend caudally to the level of the lateral cuneate nucleus in the lateral wall of the caudal portion of the fourth ventricle. The four nuclei are the rostral, medial, lateral, and caudal vestibular nuclei. They form a continuous column on each side of the pons and medulla. The rostral vestibular nucleus is located medial to the rostral and middle cerebellar peduncles, dorsal to the motor nucleus of the trigeminal nerve in the pons (see Fig. 2-11). The medial and lateral vestibular nuclei are located ventromedial to the confluence of the three cerebellar peduncles with the cerebellum (see Fig. 2-12). They are dorsal to the ventrolateral projection of the facial neurons. The medial nucleus continues caudally adjacent to the caudal vestibular nucleus in the dorsal medulla to the level of the lateral cuneate nucleus (see Fig. 2-13). The lateral vestibular nucleus is only located at the level of the confluent cerebellar peduncles (see Fig. 2-12). The caudal vestibular nucleus is caudal to the lateral vestibular nucleus and continues caudally to the level of the lateral cuneate nucleus. The caudal cerebellar peduncle is dorsolateral to the caudal vestibular nucleus. The spinal tract of the trigeminal nerve and its nucleus are ventrolateral to the caudal vestibular nucleus in the medulla. These vestibular nuclei receive afferents from the vestibular division of the vestibulocochlear nerve. From the vestibular nuclei are numerous projections, which can be grouped into spinal cord, brainstem, and cerebellar pathways (see Fig. 12-4).
Spinal Cord
The lateral vestibulospinal tract courses caudally in the ipsilateral ventral funiculus through the entire spinal cord. Its axons terminate in all of the spinal cord segments on interneurons in the ventral gray columns (see Fig. 2-17). These interneurons are facilitory to ipsilateral alpha and gamma motor neurons to extensor muscles, inhibitory to the ipsilateral alpha motor neurons to flexor muscles, and some interneurons cross to the opposite ventral gray column where they are inhibitory to the contralateral alpha and gamma motor neurons to extensor muscles (see Fig. 12-2). Thus the effect of stimulation of the neuronal cell bodies, the axons of which are in the vestibulospinal tract, is an ipsilateral extensor tonus and contralateral inhibition of this mechanism. The cell bodies of most of the axons in the lateral vestibulospinal tract are located in the lateral vestibular nucleus.
The medial vestibulospinal tract arises from cell bodies in the rostral, medial, and caudal vestibular nuclei and passes caudally in the ipsilateral ventral funiculus of the cervical and cranial thoracic spinal cord segments.49 These axons terminate on interneurons in the ventral gray columns, which influence the activation of the alpha and gamma motor neurons that innervate primarily neck muscles. In addition, the medial vestibular nucleus projects axons into the medial longitudinal fasciculus, which courses caudally in the dorsal portion of the ventral funiculus through the cervical and cranial thoracic spinal cord segments.35,36
Brainstem
Neuronal cell bodies in the vestibular nuclei have three general terminations in the brainstem.
Cerebellum
Axons of neuronal cell bodies in the vestibular nuclei, in addition to some in the vestibular ganglia, project to the cerebellum through the caudal cerebellar peduncle and terminate mostly in the cortex of the flocculus of the hemisphere and the nodulus of the vermis (the flocculonodular lobe). These axons have collaterals that synapse in the fastigial nucleus, which is the most medial of the three nuclei in the cerebellar medulla (see Figs. 12-2 and Fig. 2-13).
CLINICAL SIGNS OF VESTIBULAR SYSTEM DISEASE
Unilateral Peripheral Vestibular Disease
Normal Nystagmus
Nystagmus is an involuntary rhythmic oscillation of the eyes. Eye movements that are equal in each direction indicate a pendular nystagmus, which is uncommon, usually benign, and is associated with congenital visual system pathway abnormalities. Eye movements that are unequal, with a slow movement (slow phase) in one direction and a fast return (quick phase) of the eye to its starting position, indicate a jerk nystagmus, which can be normal or abnormal and reflect a dysfunction in the vestibular system. The direction of the nystagmus, by convention, is ascribed to the direction of the quick or fast phase of the jerk nystagmus. Both eyes are usually affected and usually in the same direction. This jerk nystagmus is a normal response to any rapid movement of the head. Stand over any normal dog and watch its eyes as you move the head in a horizontal-dorsal plane form side to side. You will observe a horizontal jerk nystagmus. As you move the head to the right, both eyes will repeatedly jerk quickly to the right with a slow return to the left. As you move the head to the left, the opposite will happen; both eyes will repeatedly jerk quickly to the left and slowly return to the right. This procedure is termed normal vestibular or physiologic nystagmus. Some textbooks refer to this response as vestibular-ocular nystagmus, or a doll’s eye response. It evaluates not only the vestibular system, which is the sensory arm of this response, but also the medial longitudinal fasciculus in the brainstem and the abducent nerve innervation of the lateral rectus muscle that abducts the eye and the oculomotor nerve innervation of the medial rectus muscle that adducts the eye. If you flex and extend the neck so that the head moves up and down, the same eye movements will occur in a vertical direction. This event is a vertical jerk nystagmus. The quick phase of the nystagmus is always in the direction of the head movement. This response is a normal reflex in which the slow component is initiated by way of the vestibular receptors in the membranous labyrinth and the quick component involves a brainstem center related to the vestibular system. This reflex is important in maintaining visual fixation on stationary points as the body rotates.
Abnormal Nystagmus
Normal nystagmus requires normal function of the vestibular system components, normal medial longitudinal fasciculus bilaterally, and normal general somatic efferent (GSE) neurons in the abducent, trochlear, and oculomotor nuclei. Abnormal nystagmus indicates a disruption in the normal bilateral balance of sensory information from the peripheral vestibular receptor and the activity of the central components of the vestibular system. No normal or abnormal nystagmus can occur with bilateral loss of function in the peripheral vestibular system, its central components, the medial longitudinal fasciculus, or the GSE motor neurons of the abducent, trochlear, and oculomotor nuclei. Bilateral otitis interna is the most common cause of the complete absence of any normal or abnormal nystagmus.
Strabismus
Strabismus is an abnormal position of the eye relative to the orbit or palpebral fissure that is a clinical sign of loss of innervation to the extraocular muscles and was described with the cranial nerves in Chapter 6. This strabismus is visible in all positions of the head. In the normal small animal, when the head and neck are extended in the tonic neck reaction, the eyes should elevate and remain in the center of the palpebral fissures. With disorders of any component of the vestibular system, this effect may not occur on the side of the lesion, resulting in a dropped or ventrally deviated eye that exposes the sclera dorsally. Occasionally, a slight ventral or ventrolateral strabismus is observed without head and neck extension but disappears when the head position is changed. This action will mimic an oculomotor nerve strabismus. However, when you move the head side to side to test for normal physiologic nystagmus, the affected eye will adduct and abduct well, indicating that cranial nerves III and VI are not impaired. This inconstant abnormal eye position is known as vestibular strabismus. You should look for this impairment when you hold the head and neck in extension because it may be the only clinical sign observed in mild disorders of the vestibular system. This vestibular strabismus will be on the same side as the lesion in the vestibular system.
Postural Reactions
The vestibular system is the only system involved with movement of the animal that, when deficient, does not interfere with the performance of the postural reactions. Hopping, hemiwalking, placing, and paw or hoof replacement will all be normal. Only the animal’s ability to right itself from a recumbent position may be altered, and this action toward the side of the lesion may be exaggerated. In the worst situation, the patient may continually roll in that direction. The ability to perform these postural reactions (except for righting) is critical to determining whether the vestibular system disorder involves the peripheral or central components of the vestibular system. You need to repeat the hopping responses many times to be comfortable that they are normal in your patient with peripheral vestibular disease. In patients with severe loss of their ability to balance, holding them securely to perform these postural reactions may be difficult. With an acute onset of severe loss of balance, delaying or repeating this part of the neurologic examination after 24 hours may be necessary to allow time for the most severe clinical signs to abate enough so that you can handle the patient for this examination. The ground surface must not be slippery and should provide good traction for the patient. Be careful if you pick up one of these patients because severe disorientation will be initiated, and they will thrash their limbs to seek a supporting surface. If you suddenly pick up a cat with this disorder, you are in danger of being grasped by the struggling patient.
Central Vestibular System Disease
Paradoxical Vestibular System Disease
Paradoxical vestibular system disease is a unique syndrome in which the head tilt and loss of balance are directed toward the side opposite to the central lesion, which usually involves the caudal cerebellar peduncle. An explanation for this paradox in the direction of the clinical signs of vestibular system dysfunction is based on the rule that the direction of the head tilt and balance loss will be toward the side of the least vestibular system activity. When we describe the physiologic anatomy of the cerebellum in Chapter 13, you will learn that the Purkinje neurons that form a single layer of cells in the cerebellar cortex are the only neurons that project their axons from the cerebellar cortex. These neurons are all inhibitory neurons that release gamma-amino butyric acid at their telodendria. Most of these neurons terminate via their telodendria on neuronal cell bodies in the cerebellar nuclei, which are located in the central portion of the cerebellum known as the cerebellar medulla. The neurons in these cerebellar nuclei comprise the majority of the efferent axons that leave the cerebellum to terminate in various brainstem nuclei. An exception to this rule is a small population of Purkinje neurons, most of which are located in the cortex of the folia of the flocculus in the hemisphere and the nodulus in the vermis. The Purkinje neurons of these cortical areas have axons that leave the cerebellum directly as a component of the caudal cerebellar peduncle. They terminate in the vestibular nuclei, where they are inhibitory to the activation of these neuronal cell bodies. A lesion in the caudal cerebellar peduncle interferes with this inhibition, resulting in excessive discharge of vestibular system neurons on that side. The imbalance in vestibular system activation between the two sides is recognized as a head tilt and loss of balance to the side opposite to this lesion because, as a rule, the direction of the head tilt and balance loss will be towards the side with the least activity of the vestibular system. This paradoxical syndrome is in contrast to lesions that cause a loss of activation of the neuronal cell bodies in the vestibular nuclei as seen in disorders of the peripheral components of the vestibular system or within the vestibular nuclei themselves.
Experimental studies support our clinical observations and proposed explanation.25 Ablation of the caudal cerebellar peduncle dorsal to the medulla on one side will produce a head tilt and balance loss directed toward the side opposite to the lesion with the nystagmus directed toward the side of the lesion. If the vestibular nuclei are included in this lesion, the head tilt and balance loss will be directed toward the side of the lesion, and the nystagmus will be toward the side opposite to the lesion, similar to disorders of the vestibular nerve or its receptors. Similarly, ablation of the flocculus and nodulus within the cerebellum will produce this paradoxical vestibular system syndrome, with the clinical signs directed toward the side opposite to this cerebellar lesion. However, experimental ablation of the fastigial nucleus, a source of activation of the vestibular nuclei, causes ipsilateral vestibular system signs.



