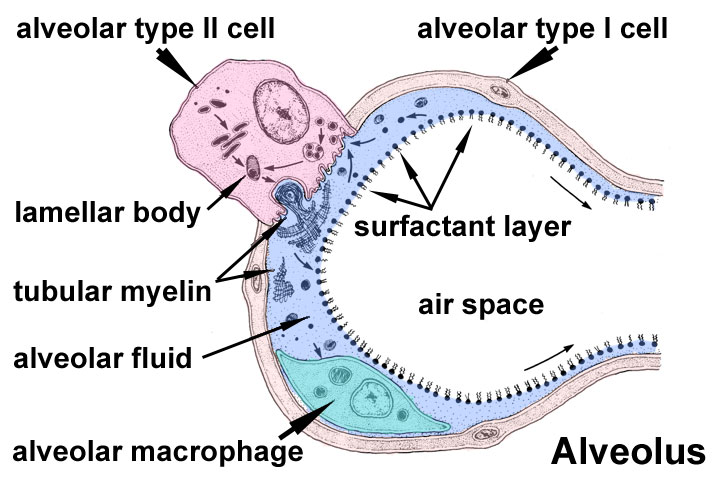
As animals evolved from single‐celled organisms to multilayered organisms, it became impossible for oxygen to diffuse effectively to all cells. Consequently, systems evolved to deliver oxygen and nutrients and eliminate waste products. The development of the respiratory and circulatory systems solved these problems. The functions of the respiratory system include gas exchange, regulating blood pH, olfaction, filtering inspired air, producing sounds, and elimination of water and heat via expired air. The primary function of the respiratory system is the intake of oxygen and elimination of carbon dioxide from the body. The respiratory system consists of the nose, pharynx, larynx, trachea, bronchi, and lungs. Structurally, the respiratory system can be divided into the upper respiratory system, including the nose, pharynx, and associated structures, and the lower respiratory system, which includes the larynx, trachea, bronchi, and lungs. Functionally, there are two sections: (1) the conducting portion, which consists of a series of connected tubes, both outside and within the lungs, that filter, warm, moisten, and conduct air to and from the lungs. This portion includes the nose, pharynx, larynx, trachea, bronchi, bronchioles, and terminal bronchioles and (2) the respiratory portion, the site of gas exchange between the air and blood, which consists of the respiratory bronchioles, alveolar ducts, alveolar sacs, and alveoli. The nose is the externally visible portion of the respiratory system. It can be divided into an external and internal portion. The external nose consists of the bone, hyaline cartilage, muscle, skin, and mucous membrane protruding from the face. The immovable portion of the external nose consists of the rostral ends of the nasal bones and the incisive bones. The nasal cartilage extends rostrally from these bones. The external nares (nostrils) are the external openings to the respiratory tract (Fig. 14.1). The philtrum is the area between the lips and nose. It is relatively deep in carnivores and small ruminants but shallow or absent in pigs, oxen, and horses. The pig possesses a rostral bone in the tip of its flattened, cylindrical‐shaped nose, apparently to assist in rooting. The lateral portion of the nose has sebaceous and sweat glands. The most rostral portion of the nose lacks sebaceous glands, except in the horse. The nasal cavity extends from the external nares to the caudal nares, and it is separated from the mouth by the hard and soft palates (Fig. 14.2). The hard palate consists of the horizontal portions of the incisive, palatine, and maxillary bones; the soft palate is a musculomucosal extension of the hard palate dividing the rostral part of the pharynx into the oropharynx and nasopharynx. The space inside the internal nose is the nasal cavity, which is divided into two halves by the median nasal septum. This septum is made of the vomer, nasal, and ethmoid bones, as well as cartilage. The nasal cavity is divided into three sections. The vestibule is the most rostral portion, located just inside the nostrils. The middle section is filled with the nasal conchae, which are scrolls of bone arising from the lateral wall and are covered with a mucous membrane (Fig. 14.2). Named superior to inferior, the three conchae are dorsal, ethmoidal (middle), and ventral nasal concha. The areas between the conchae are called meatuses and include the dorsal, middle, and ventral nasal meatus. The common nasal meatus is located between the median nasal septum and conchae, and it is continuous with the other nasal meatuses. Fig. 14.1 External nose of cat (left) and dog (right). Used with permission from Constantinescu, G.M., 2002. Clinical Anatomy for Small Animal Practitioners. Iowa State Press, Ames, Iowa. Fig. 14.2 Median aspect of the head of a large ruminant. The dorsal, ventral, and middle nasal concha are visible in the nasal cavity. They are located dorsal to the hard palate and are separated by dorsal, middle, and ventral nasal meatuses. Also shown are the oropharynx, nasopharynx, and laryngopharynx. Reprinted from Constantinescu and Constantinescu (2004). Used by permission of the publisher. The caudal section of the nasal cavity contains many ethmoturbinates (conchae of the ethmoid bone). The nasal cavity communicates with the paranasal sinuses and posteriorly with the nasopharynx through two openings called the internal nares, or choanae (Fig. 14.3). The paranasal sinuses are air‐filled cavities within some bones of the skull. The major ones are the frontal and maxillary sinuses, but others may be present depending on the species. In horned cattle, the frontal sinus can extend into the horn as the cornual diverticulum. Fig. 14.3 Paranasal sinuses. (A) Lateral aspect of a horse. (B) Frontal aspect of a horse. (C) Lateral aspect of a large ruminant. (D) Frontal aspect of a large ruminant. Reprinted from Constantinescu and Constantinescu (2004). Used by permission of the publisher. The pharynx connects the nasal cavity and mouth to the larynx and esophagus, respectively. Commonly called the throat, it directs food and air into the digestive and respiratory systems, respectively. The soft palate divides the rostral portion of the pharynx into the oropharynx and nasopharynx (Fig. 14.2), and the common caudal portion is the laryngopharynx. The nasopharynx is located dorsal to the soft palate, extending from the internal nares to the laryngopharynx. The palatopharyngeal arches are found at the border of the nasopharynx and laryngopharynx. Because it is located above the mouth, it serves as a passageway only for air. When the animal swallows, the soft palate and uvula move superiorly, closing off the nasopharynx and preventing food and water from entering the nasal cavity. The pharyngotympanic, or auditory, tubes drain from the middle ear to the nasopharynx. It helps equalize the pressure within the middle ear with atmospheric pressure. The oropharynx lies ventral to the soft palate, extending from the oral cavity to the base of the epiglottis. The palatoglossal arches lie at the border between the oral cavity and the oropharynx. This is a common pathway for both swallowed food and inhaled air. The laryngopharynx also serves as a common pathway for food and air. It extends from the epiglottis to the larynx, the diverging point for the respiratory and digestive systems. The larynx connects the laryngopharynx with the trachea and contains the vocal cords. The two functions of the larynx are: (1) provide a routing mechanism for air and food and (2) make sounds. Superiorly, the larynx attaches to the hyoid bone. The larynx is formed by five mucous‐covered cartilages, including single epiglottic, thyroid, and cricoid cartilages, as well as paired arytenoid cartilages (Box 14.1). The epiglottic cartilage provides structure to the epiglottis, which closes the opening to the larynx during swallowing, thus preventing ingested materials from entering the lungs. The thyroid cartilage is the largest cartilage and forms the “Adam’s apple” in humans. The cricoid cartilage connects the thyroid cartilage and trachea. The arytenoid cartilages are paired and irregularly shaped. They have a ventral vocal process to which the vocal ligament (vocal cord) is attached. The glottis consists of the vocal ligaments and the slit‐like gap between them, the glottic cleft. The trachea, or windpipe, is a cylindrical tube extending from the larynx to the bifurcating right and left primary bronchi above the base of the heart. The cervical portion runs from the larynx to the thoracic inlet, while the thoracic portion continues to the bifurcation of the primary bronchi. The thoracic inlet is formed by the first pair of ribs, the first thoracic vertebra, and the cranial parts of the sternum. The trachea consists of four layers: (1) mucosa, which is the deepest layer; (2) submucosa; (3) hyaline cartilage; and (4) adventitia, the most superficial layer composed of areolar connective tissue. The mucosa is pseudostratified ciliated columnar epithelium resting on the lamina propria containing elastic and reticular fibers. The trachea contains a series of dorsally incomplete, C‐shaped hyaline cartilage rings. These rings keep the trachea open. The tracheal cartilages are united by the annular cartilage, which makes the trachea flexible (Fig. 14.4). The trachealis muscle is a smooth muscle connecting the open, dorsal portion of the cartilaginous rings. The trachea divides into the right and left primary, or principal, bronchi. Such as the trachea, the primary bronchi contain incomplete cartilaginous rings and are lined with pseudostratified ciliated columnar epithelium. Upon entering the lungs at the lung’s hilus, the primary bronchi divide into the smaller secondary, or lobar, bronchi. These keep dividing into the following sequence of tubes: tertiary, or segmental, bronchi > bronchioles > respiratory bronchioles > alveolar ducts > alveolar sac > alveoli (see Fig. 14.5; see also Fig. 14.7). This extensive branching of the respiratory tubes is called the bronchial tree. The alveoli are thin‐walled sacs where gas exchange occurs. Below the tertiary bronchi, the mucous membrane changes from ciliated cuboidal epithelium with goblet cells to nonciliated simple cuboidal epithelium in the respiratory bronchioles. Fig. 14.4 Trachea of a goat. (A) Cross section of trachea. (B) Annular ligaments. Reprinted from Constantinescu (2001). Used by permission of the publisher. Fig. 14.5 The canine bronchial tree. Reprinted from Constantinescu (2002). Used by permission of the publisher. The lungs are paired organs located within the thorax. In general, the left and right lungs have two and four lobes, respectively. The horse has three right lobes (Fig. 14.6). The cranial portion, or apex, of each lung is in the thoracic inlet; the base is the caudal end of the lung resting on the diaphragm. The hilus of the lung is the medial area where the bronchi, blood vessels, and nerves enter the lungs. The cardiac notch is the indentation between the lobes where the heart contacts the lung. The lungs are surrounded by a serous membrane called the pleural membrane. The superficial layer lining the thoracic cavity is the parietal pleura, and the layer closely adhering to the lungs is the visceral pleura. The narrow parietal space between these two layers contains a small amount of pleural fluid that allows the two layers to slide over one another during breathing. Inflammation of the pleural membrane is called pleurisy. The mediastinum is the midline site formed where the two pleural membranes meet. It contains the heart, large vessels, esophagus, and other structures, and separates the two lungs from one another. After the first breath, the lungs become less dense. This fact allows one to determine whether a newborn animal is stillborn. A sample of lung tissue can be placed in water to see whether it floats. If it floats, it indicates that the animal took at least one breath and therefore was born alive. The pressure inside the intrapleural space is negative. This negative pressure is vital for the expansion of the lungs. If an injury to the chest wall punctures the pleural membrane, it can allow air to enter the intrapleural space, resulting in a pneumothorax. Such an injury can be caused by a sharp object penetrating the chest cavity or a traumatic blow such as being struck by a car. Because this allows the intrapleural pressure to equilibrate with atmospheric pressure, the lung on that side will collapse. Surrounding the alveolar ducts are many alveoli and alveolar sacs. Within the alveolar sac are two or more alveoli, balloon‐like bulges, lined with simple squamous epithelium resting on a thin elastic basement membrane (Fig. 14.7). Alveoli walls contain predominantly type I alveolar cells, which are simple squamous epithelial cells. These are the primary sites of gas exchange. They also contain type II alveolar (or septal) cells, alveolar macrophages, and fibroblasts that produce reticular and elastic fibers. Type II alveolar cells are cuboidal epithelial cells containing microvilli that secrete alveolar fluid containing surfactant. The alveolar macrophages are wandering phagocytes that remove debris from the lungs. Fig. 14.6 Lungs of a horse. (A) Medial aspect of the left lung. (B) Medial aspect of the right lung. Reprinted from Constantinescu and Constantinescu (2004). Used by permission of the publisher. Fig. 14.7 Alveolar sacs and respiratory membrane. Within the alveolar sac are several alveoli. The alveoli contain type I alveolar cells that are the site of gas exchange. Type II alveolar cells produce surfactant. Adapted from http://php.med.unsw.edu.au/embryology/images/d/d4/Alveolar‐sac‐01.jpg. The respiratory membrane is where O2 and CO2 diffuse across the alveolar and capillary walls. It is a very thin membrane about 0.5 μm thick and consists of four layers. The pulmonary and bronchial arteries supply blood to the lungs. The pulmonary artery carries deoxygenated blood through the pulmonary trunk and into the right and left pulmonary arteries. Oxygenated blood returns to the left atrium via the pulmonary veins. In most vessels, hypoxia causes dilation of blood vessels to increase O2 delivery. However, in the pulmonary blood vessels, hypoxia causes vasoconstriction. This is called ventilation–perfusion coupling, which allows pulmonary blood to be diverted to well‐ventilated areas. Bronchial arteries arise from the aorta and deliver oxygen to the lungs. They mostly perfuse the bronchi and bronchioles. Their blood returns mainly via the pulmonary veins, with some return via the superior vena cava. During exercise, cardiac output can increase by as much as eightfold. During this time, blood flow to the lungs must also increase to collect oxygen. Pulmonary blood vessels dilate in response. In the horse, pulmonary arterial pressure can be high enough to cause erythrocytes to leak from pulmonary capillaries, a condition called exercise‐induced pulmonary hemorrhage. This is a significant problem in athletic horses. Respiration, the process of gas exchange, occurs in three steps: Boyle’s law states that at constant temperature and pressure, there is an inverse relationship between the volume and pressure of a gas: where P is the pressure of the gas (mL of Hg), and V is the volume (cm3). Therefore, the pressure inside a closed container will decrease as the volume of the container increases, and vice versa. When this law is applied to the lungs, we find that the pressure inside the lungs decreases as the volume of the lungs increases. Air moves into the lungs because of a decrease in air pressure within the lungs. In most animals, inspiration is an active process. The main muscles responsible for quiet inhalation are the diaphragm and external intercostal muscles. The diaphragm is a dome‐shaped muscle innervated by the phrenic nerves. As the diaphragm contracts, it increases the horizontal dimensions of the thoracic cavity, thus increasing the volume of the thorax. This causes the volume of the lungs to expand, causing a decreased pressure inside the lungs. This results in inspiration. Contraction of the external intercostal muscles causes the ribs to move cranially and ventrally, which also increases the diameter of the thorax. This change accounts for about 25% of the entry of air into the lungs with each inspiration cycle. In a horse at rest, the intrapleural pressure is negative, about 754 mmHg. As inspiration begins, this pressure drops to approximately 744 mmHg. Atmospheric pressure is 760 mmHg at sea level. As the volume of the thoracic cavity increases during inspiration, the parietal pleura is pulled outward, and the visceral pleura is pulled with it. As a result, the pressure inside the lungs, the alveolar pressure, decreases. The air then flows from an area of high pressure (the atmosphere) to an area of lower pressure (the alveoli). As an animal increases the force of inspiration, additional muscles are engaged. These include the sternocleidomastoid muscles that move the sternum rostrally, the scalene muscles that pull the first two ribs forward, and the pectoralis minor muscles that pull several other ribs forward. Normal expiration is a passive process involving no active muscle contraction. Like inspiration, it is due to pressure gradients, but in the opposite direction. Because of the elastic recoil of the lungs and chest wall, there are two inwardly directed forces producing this recoil: (1) the elastic fibers that were stretched during inhalation and (2) the inwardly directed force due to the surface tension arising from the alveolar fluid. As the neural signals to the diaphragm cease, it relaxes, and this dome‐shaped muscle moves rostrally, thus decreasing the volume of the thoracic cavity. The external intercostal muscles also relax, allowing the ribs to move dorsally and caudally, thus further decreasing the volume of the thoracic cavity. This decreases lung volume and causes alveolar pressure to increase approximately 2 mmHg above atmospheric pressure. As a result, air flows out of the lungs to an area of lower pressure. During forceful exhalation, the abdominal and internal intercostal muscles contract. This causes the ribs to move caudally and dorsally, compressing the abdominal viscera and decreasing the thoracic volume. This increases pressure inside the thoracic cavity and forces air outward. Alveolar fluid coats the inside surface of the alveoli. Because of hydrogen bonding, this fluid has a surface tension. In a sphere such as that found in the alveoli, this surface tension produces an inwardly directed force causing the alveoli to assume the smallest possible diameter. To expand the lungs, this surface tension must be overcome. This surface tension accounts for approximately two‐thirds of the lung’s elastic recoil. Surfactant, produced by type II alveolar cells, is a complex of lipids and proteins that reduces the surface tension in much the same way that soap allows lipids to dissolve in aqueous solutions. Because surfactant is one of the last compounds produced during embryonic development, premature animals often have respiratory distress because of the underdeveloped respiratory system. In the case of sheep, surfactant is released into the alveolar spaces near the beginning of the fourth month of gestation. Its release correlates with a rise in plasma cortisol levels (Box 14.2). The distensibility of the lungs is referred to as lung compliance. High lung compliance means that the lungs will expand easily. Lung compliance is related to (1) the distensibility of lung tissue and the thoracic cage and (2) alveolar surface tension. Lung compliance is normally high due to the elasticity of the lung’s tissue and decreased alveolar surface tension because of surfactant secretion. Compliance can be decreased by several factors: (1) scar tissue formed in the lungs because of certain diseases, (2) pulmonary edema resulting from the accumulation of fluid in the lungs, (3) insufficiency of surfactant, and (4) decrease in the ability of the thoracic cage to expand. The flow of air into the lungs is inversely related to airway resistance: where F is the gas flow, P is the pressure, and R is the resistance. The walls of the airways into the alveoli, particularly the bronchioles, create resistance to airflow. The larger the diameter of the airway, the less the resistance to airflow. The diameter of the airways can be altered by the degree of contraction of the smooth muscle in these airways. Stimulation of the sympathetic nervous system causes relaxation of these walls, which allows air to more readily enter the lungs. Diseases or injuries to the airways can increase airway resistance. During exercise, animals decrease airway resistance by dilating the external nares and vasoconstriction of the vascular tissue in the nose. As exercise rate increases, cows and dogs breathe through their mouth to bypass the greater airway resistance associated with the nose. However, horses are considered obligatory nose breathers and must rely on their ability to decrease airway resistance to increase airflow. One can watch the nostrils of a horse flare during exercise (Box 14.3). The respiratory capacities, or the amount of air that moves in and out of the lungs, depend on the strength of inspirations and expirations. During normal, quiet breathing, the volume of air moving in and out of the lungs is called the tidal volume (TV). Only about 70% of TV reaches the lungs. The remaining portion of the air is found in the airways, including the nose, pharynx, larynx, trachea, bronchi, bronchioles, and terminal bronchioles. These airways are collectively called the anatomical dead space. The minute volume (MV) is the volume of air inhaled and exhaled each minute and is calculated as: Table 14.1 Tidal volume and respiration rate.a Fig. 14.8 Respiratory volumes and capacities. During quiet respiration, the volume of air inspired and expired is called the tidal volume. The additional air brought into the lungs during a forced inspiration is called the inspiratory reserve volume, and the additional air that can be expired during a forced expiration is called the expiratory reserve volume. The air remaining in the lungs after a forced expiration is called the residual volume. The respiration rate varies by species (Table 14.1). Because of the anatomical dead space, not all the MV is available for gas exchange. The alveolar ventilation rate (AVR) is that portion of the TV that reaches the site of gas exchange: If an animal inhales more forcefully, it can increase the volume of air entering the lungs above normal TV (Fig. 14.8). The additional inhaled air is called the inspiratory reserve volume. Similarly, an animal can force more air out of its lungs than occurs during quiet respiration. This additional exhaled volume is called the expiratory reserve volume. Following a forced expiration, the air remaining in the lungs is the residual volume. The exchange of oxygen and carbon dioxide between alveolar air and pulmonary blood is a passive process, explained by two gas laws. Dalton’s law describes how gases move by diffusion based on pressure differences; Henry’s law describes the diffusion of gas based on its solubility. Dalton’s law states that each gas within a mixture exerts its own pressure independent of the other gases present. The pressure of an individual gas is called its partial pressure, designated Px. The total pressure of a gas mixture is calculated by summing all its partial pressures. Atmospheric air is primarily a combination of nitrogen (N2), oxygen (O2), carbon dioxide (CO2), and water vapor. At sea level, atmospheric pressure is 760 mmHg. Atmospheric air is 78.6% nitrogen, 20.9% oxygen, 0.04% carbon dioxide, 0.06% other gases, and varying water vapor, depending on the humidity. Therefore, atmospheric air pressure can be computed as follows:
14
Respiratory System
Introduction
Functional Anatomy: Nose and Paranasal Sinuses



Conducting Pathway Anatomy
Pharynx and Nasopharynx
Oropharynx and Laryngopharynx
Larynx
Trachea and Bronchi


Respiratory Anatomy of the Lungs and Pleural Membrane
Alveoli


Blood Supply to the Lungs
Pulmonary Ventilation
Pressure Changes During Respiration
Inspiration

Expiration
Other Factors Involved in Pulmonary Ventilation
Surface Tension of Alveolar Fluid
Compliance of the Lungs
Airway Resistance

Lung Volumes and Capacities

Species
Body Weight (kg)
Condition
Respiration Rate (Breaths/min)
Tidal Volume (mL/kg Body Weight)
Cat
3.7
Anesthetized
30
9.2
Holstein cow
516
Standing
26
8.2
Jersey cow
450
Standing
27
8.4
Dog
19
Anesthetized
13.6
10.7
12.6
Anesthetized
21.0
11.4
Horse
486
Resting
10
15.4


Exchange of Oxygen and Carbon Dioxide
Dalton’s Law and Henry’s Law
Dalton’s Law




Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree



{kind=link}