CHAPTER 118 Ovarian Function in Llamas and Alpacas
South American camelids possess some unique reproductive characteristics generally unfamiliar to veterinarians practicing outside the indigenous zone for camelids, the Andes mountains. Camelids are the only large domestic species that are induced ovulators; females are in virtually constant estrus until successful mating induces ovulation. The term estrous cycle must be qualified when used in reference to induced ovulators because a regular cyclic pattern of behavior does not occur as it does in spontaneous ovulators. The terms receptive versus nonreceptive, and follicular phase versus luteal phase are more appropriate for communicating “cyclic” changes in camelids. Sexual behavior and copulation time is strikingly different from any other domestic species, gestation is unusually long, and uterine anatomy, placentation, and birthing all have distinctly different characteristics in camelids than any other species. Although the reproductive characteristics of llamas and alpacas appear to be virtually identical, extremely little work has been done directly comparing the two species. As veterinarians, we have been frustrated at the lack of information about the normal reproductive function of camelids. Our inability to distinguish normal from abnormal has made it difficult to diagnose or prognose clinical conditions, or make any meaningful recommendations regarding breeding management. This chapter is intended to focus on the basic aspects of reproductive physiology of llamas and alpacas. Emphasis is placed on ovarian function including follicle and luteal dynamics, and the implications of endogenous patterns on clinical diagnosis, treatment, and reproductive management.
FOLLICULAR DYNAMICS
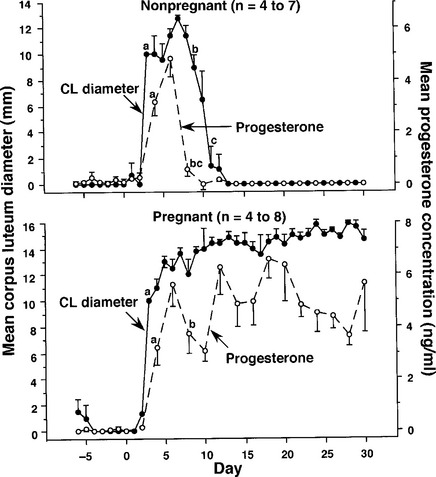
As induced ovulators, three naturally occurring reproductive statuses exist in llamas and alpacas: (1) nonovulatory, (2) ovulatory but not pregnant, and (3) pregnant. In a study involving ultrasonographic examination of llamas (n = 41; Fig. 118-1) daily for a period of 60 days or longer,1 ovarian follicle development was found to follow a wave-like pattern regardless of reproductive status (nonovulatory, ovulatory nonpregnant, or pregnant) or lactational status (lactating, nonlactating). That is, a group of follicles begins to grow synchronously, one of which continues to grow and become dominant while the others (subordinates) grow for a short period and then regress. If ovulation is not induced, the dominant follicle eventually regresses as well, and a new wave emerges so that the ovarian “cycle” repeats itself (see Fig. 118-1). The interval between emergence of successive waves of follicles was longer in nonpregnant animals (20 days) than in pregnant animals (15 days), and lactation was associated with a 2.5-day abbreviation in the interwave interval. Maximum diameter of nonovulatory dominant follicles ranged from 9 to 16 mm and was greater, on average, in nonpregnant animals (12 mm) than in pregnant animals (10 mm). Dominant follicles of successive waves are equally as likely to develop in the ipsilateral as contralateral ovary; they do not regularly alternate between ovaries.1,2
In a recent ultrasonographic study, the wave pattern of follicle development was also documented in alpacas.2 The mean (±SEM) interwave interval was 15.4 ± 0.5 days and ranged from 12 to 22 days. Individual alpacas exhibited relatively short as well as longer interwave intervals, and body weight did not appear to be related to the length of the interval. However, the growth rate of dominant follicles was consistent over the first 10 days after emergence and the dominant follicle reached a diameter capable of ovulation by this time, regardless of subsequent interwave interval. The authors concluded that the optimal time of mating might be predicted in alpacas, provided that the emergence of ovarian follicular waves was controlled.
OVULATION
Female llamas and alpacas ovulate only after copulation, or after the administration of hormones with LH (luteinizing hormone)-like activity.3,4 In an early study designed to determine factors associated with eliciting ovulation,4 it was concluded that mounting with penile intromission is necessary to induce ovulation, regardless of whether the male is intact or vasectomized. Neural stimuli from penile intromission, treading and clasping of the male’s legs on the back and sides of the female, and gutteral humming sounds emitted by the male are thought to influence ovulation. Circulating concentrations of LH start to rise 15 minutes after the start of copulation, peak at approximately 2 to 4 hours, and decrease to basal values by 6 hours after copulation.5,6 Based on daily ultrasonography,1,7 ovulation occurred with remarkable consistency (96% of the llamas) by the second day after the first of two matings (4 to 8 hours apart) with either a vasectomized or intact male. The mean diameter of the ovulatory follicle on the day before ovulation was 10 mm and it ranged from 7 to 14 mm. In a more recent study involving ultrasonography every 4 hours,8 the interval from mating to ovulation was 30.0 ± 0.5 hours (mean ± SEM), and was not different from the interval to ovulation following gonadotropin-releasing hormone (GnRH) or LH treatment.
Interestingly, it appears that a unique ovulation-inducing factor (OIF) is present in the seminal plasma of camelids. In a preliminary study,9 alpacas were selected (n = 58) when a growing follicle of 8 mm or more in diameter was detected, and were assigned to groups treated with seminal plasma or saline intramuscularly or by intrauterine infusion. Ovulation was detected only in alpacas given seminal plasma intramuscularly (13 of 14, 93%). It was concluded that alpaca seminal plasma contains an OIF that acts via a systemic route. Bactrian camels are the only other species in which an OIF has been reported.10 The importance of OIF relative to physical stimuli for inducing ovulation during natural mating awaits further investigation.
Unmated females remain in a follicular phase characterized by more or less constant sexual receptivity,11,12 but sexual behavior does not appear to be related to follicle size because females are sexually receptive with follicle diameters ranging from 5 to 12 mm.13 However, follicle size is related to circulating estrogen concentrations5 and ovulatory responsiveness.14 Females with small follicles (4–5 mm) did not ovulate after copulation, whereas females with follicles 7 mm and larger and in the growing phase, or females with mature follicles (8–12 mm) did ovulate after copulation. Regressing dominant follicles did not ovulate after copulation or LH administration.
Even in the presence of a mature follicle, ovulation does not always occur after copulation. Ovulation failure occurs in approximately 20% of pasture-mated females15 and 10% of hand-mated females.6 This may be due to inadequate amounts of LH released by the pituitary gland. In this regard, it has been reported that a second copulatory period at 6, 24, or even 48 hours after the first copulatory period did not invoke more release of LH.6 Correspondingly, an injection of GnRH at 6, 24, or 48 hours after copulation did not invoke more LH release.6 It appears that if ovulation is to occur at all, it will occur subsequent to the first copulatory stimulus or GnRH/LH administration. The incidence of spontaneous ovulation has been reported to be 4% to 8%.4,7,16 In some instances, spontaneous ovulation may be associated with the presence of males, owing to visual, olfactory, and auditory stimuli.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


