(1)
Indian Institute of Science Education and Research Pune (IISER-P), Pune, India
Abstract
I will make a fresh beginning now that does not involve any destructive criticism but instead tries to synthesize a new alternative picture that is logically coherent as well as supported by evidence. This necessitates leaving the thread of diabetes for the time being to which we will return shortly. We need to look at certain aspects of the life and behavior of animals in the wild. Since the human race evolved in the same wilderness, it is very likely that any principles learned there may help us give a novel insight in human life, behavior, physiology, and health.
I will make a fresh beginning now that does not involve any destructive criticism but instead tries to synthesize a new alternative picture that is logically coherent as well as supported by evidence. This necessitates leaving the thread of diabetes for the time being to which we will return shortly. We need to look at certain aspects of the life and behavior of animals in the wild. Since the human race evolved in the same wilderness, it is very likely that any principles learned there may help us give a novel insight in human life, behavior, physiology, and health.
Animals have been used to understand human health and disease for a long time. But we have so far relied almost entirely on captive animals. There are many obvious advantages of using captive animals, the main being the ability to control their environment and thereby minimizing the number of variables. However, in doing so, we are certainly missing an insight into a number of subtle issues in physiology. Wild animals are rarely used to gain insights into physiology for obvious difficulties in controlling the conditions and collecting samples for analysis. Fortunately this picture has started changing of late, at least marginally owing to increased rigor in behavioral ecology research on the one hand and better sampling and assay techniques on the other. Another upcoming trend relevant and useful for us is the insight in human behavior obtained from animal studies. For decades psychology, the science of human behavior, and ethology, the science of animal behavior, have developed parallel to each other without sufficient give and take, often deliberately avoiding any possible points of intersection. For example any reference of animal personalities still evokes strong reactions from many sectors. Personality is about consistent and intrinsic individual differences. There is increasing evidence that individual animals consistently differ in their responses to a given challenge, but still there is some reluctance to call them personality differences. It is only recently that there are some signs of the ice breaking, and there is increasing acceptance to the concept of animal personalities [1–3]. The long-standing reluctance has a historical root in the way Western and Eastern philosophies developed and science originated. In many of the Eastern philosophies, animals are not qualitatively different from humans. The differences perceived are quantitative. In Hinduism, for example, animals are believed to have a soul as much as humans. Not only that, the same soul can take human and animal forms in the natural course of the believed chain of births. The residual effect of this philosophy on the Eastern mind is that animals are viewed at par with humans albeit differences such as expression of thoughts in the form of a spoken language. In contrast in the mainstream traditional Western thought, humans are qualitatively different and are believed to have a soul which animals need not have. Mind and emotions in animals are often denied, or at least doubts are expressed about their existence. This has relevance to science and particularly to medicine. Animals are almost invariably used in research in medicine. Most of our understanding of human physiology has come from animal experiments. There is no conceptual problem about this. However, that is not the case when it comes to cognition and emotions. Few objections are raised in applying an animal experiment to infer something about human physiology. But this is not acceptable to many in the context of behavior or emotions. Since humans have evolved from animal lineages, it would be natural to assume that the principles of evolution that we learn from plant–animal observations and experiments are applicable to humans as well. But generally this is not the baseline assumption in most branches of humanities. Recently this picture appears to be changing with subjects like evolutionary psychology taking roots as a serious branch of science. But they have not enjoyed a smooth path to acceptance so far.
This debate is relevant to us because I think we can learn much about human health from animal behavior, the link going through human behavior. We need to accept that although the factors and forces shaping human behavior are not identical to those shaping animal behavior, we have evolved following the same evolutionary principles as they have. Many of the fundamental principles of animal behavior were largely applicable to human behavior through much part of our ancestry. People who object to talking about human and animal behavior on the same platform generally do not mind talking about human and animal physiology on the same platform. The parallels or in fact almost identical physiological mechanisms across different species of animals and humans are what the current paradigm accepts very well. Human physiology evolved in an era when we were not too different from animals. Therefore at least in the context of behavior–physiology interface, there should not be any conceptual hurdles about learning some fundamental principles from the animal world.
Although we will use a number of principles from behavioral ecology in understanding human health and disease, we will make a beginning with and focus on multiple behavioral strategies, the ways animals make choices out of the available strategies, and the contextually adaptive or maladaptive effects of these choices. The problem of choosing between alternative behavioral strategies is often addressed using a branch of interesting mathematics called game theory which was introduced briefly in Chap. 1. It is time to make use of that concept now.
The hawk and dove (HD) game is one of the fundamental and earliest models of evolutionary game theory addressing the problem of physical aggression [4]. There are two basic strategies, one called “hawk” which is aggressive and the other called “dove” which avoids physical aggression. A dove may use threatening displays but will retreat invariably if it comes to a physical fight. The assumptions of the game are that there is a resource for which there is some conflict. The nature of the conflict is such that one who wins gets a reward. If the conflict is between a hawk and a dove, dove will always retreat and hawk will get the reward. Therefore we may be tempted to think that it is always good to be a hawk. But this is not true, and we can realize this on looking at what happens when dove meets a dove and hawk meets a hawk. If two doves meet, they may give threat displays and try to force retreat on the other but will never launch an attack. One of the two wins this psychological war eventually and gets the reward. Alternatively, they may negotiate and share the reward. They may have to spend some time and energy in the threatening displays or negotiations, but an actual fight never ensues. On the other hand, when two hawks enter a conflict, a fight is inevitable. One of them wins the fight ultimately and gets the reward. But whenever there is a physical fight, there is a risk of serious injuries which is a great cost. The loser in a hawk–hawk interaction neither gets the reward nor is spared from the risk of injury. The benefit in a dove–dove and hawk–hawk interaction is the same, but the mean cost of injury in a hawk–hawk interaction is large. This is the major disadvantage to hawks.
We can handle this game using the concept of an evolutionarily stable strategy (ESS) now. If the population consists of all doves and if there arises a hawk by mutation or immigration, the hawk will always win and therefore will be at an all-time advantage. Therefore dove is not an ESS. On the other hand, if all are hawks in a population and a dove invades, the dove will never win a battle, but it will also never get injured, whereas hawks will keep on fighting with each other and paying a huge cost of injuries all the time. Therefore a dove might actually do better in an all-hawk population. Therefore hawk is also not an ESS. This can be shown in a payoff matrix (see Appendix III). We are making a simple assumption here that when there is a hawk–hawk encounter, each animal has an equal a priori chance of winning. The same applies to dove–dove encounters.
Since hawk gets an advantage in a dove population and dove does better in a hawk population, there is a negatively frequency-dependent selection. Such a negatively frequency-dependent selection would lead to an optimum proportion of hawks and doves where the mean payoff of hawks and doves is exactly equal. The equilibrium proportion of hawks and doves can be easily calculated by simple algebra (see Appendix III for the mathematical framework).
The algebraic solution of the problem has two possible interpretations. In one, each individual is either a pure hawk or a pure dove, and the population comprises an equilibrium proportion of the two such that the net average payoff of everyone becomes equal. In the other a single individual can behave as a hawk or a dove in an optimum ratio given by the algebra. This is called mixed strategy. A mixed strategy at an optimum proportion can be an ESS provided there is no cost associated with the ability to posses this behavioral flexibility. But if behavioral plasticity has substantial cost, then all the three can coexist, i.e., there can be pure hawks, pure doves, and mixed strategists in the population simultaneously at some equilibrium proportion.
It is very obvious that the model is too naïve as compared to the reality of animal behavior. Realities are more complex than what is assumed in the baseline model. There have been a number of attempts to bring in the complexities of real life into the theoretical model [5, 6]. We will discuss some of the complexities that are relevant to us without going into the depths of theoretical modeling. For example, the simple assumption that each animal has an equal chance of winning in a hawk–hawk encounter is certainly not true. Individuals differ in their size and strength, and a stronger animal will have a higher probability of winning a physical combat. Animals have mechanisms of judging their size and strength vis-a-vis the opponent, and these judgments will influence the behavioral decision. An individual may actually take a decision to behave as a hawk or dove based on this judgment. When faced with a larger and stronger animal, it would prove beneficial to act like a dove, and when interacting with a weaker animal, it will be most appropriate to behave like a hawk. A number of such factors certainly play a role in actual decision making. The model is not intended to apply to behavior of any species in its naïve form. Nevertheless a number of inferences of the model remain robust enough not to be altered by the subtleties and complexities of real life.
One of the important inferences of the solution of HD game is that two or more behavioral strategies can coexist in a population leading either to a genetic polymorphism or to behavioral plasticity. A more complex example of coexistence of alternative strategies is that of lizard males with small territories, those with large territories and those without territories that coexist in a rock–paper–scissor-like game [7]. The rock–paper–scissor (RPS) game is another slightly more complex game in which three strategies can coexist with a negative frequency dependence resulting into stable or oscillating dynamics.
Another important inference of the hawk and dove model is that different alternative behavioral strategies may have their own advantages under different conditions and at different frequencies. The behavior of the dove is of particular interest. Although the dove never escalates aggression and always retreats against a hawk, it does give it a try by giving a threat display. This is a deceptive behavior which works at times. This can be viewed as a kind of social smartness. If an individual is not strong enough to win a physical combat, it can still hope to gain by using tactical deception. Tactical deception is indeed observed in animals, particularly (but not restricted to) primates [8–13], and by the above logic, we expect that physically weaker individuals should display deception more frequently. Owing to the availability of an alternative strategy to gain fitness, subordinate individuals can hope to attain some share of reproductive success. As a result social dominance does not remain synonymous with Darwinian fitness.
There are many real-life examples of hawk- and dove-like strategies in animal populations. In a wide variety of species, males compete for females, and mating behavior necessitates aggression and dominance. However males that are weak, submissive, or subordinate are not complete reproductive failures. In a wide variety of species, subordinate males remain with the harem submissively and sneak mate opportunistically. Sneak mating is a fairly successful reproductive strategy suitable for physically weaker individuals [14–20]. It takes different forms in different species depending on the ecology and sexual behavior. For example, in frogs that have external fertilization, males attract females in the breading season by vocal signals. Larger frogs have large vocal sacs and can therefore attract a large number of females. Smaller frogs that cannot do this linger around larger males, and when the larger males attract females, the satellite males sneak some reproductive success [21]. Researchers claim that in order to do this, they make use of information about neighboring males in an organized fashion [21]. As a result using more than one alternative strategy is common even in these, believed to be less intelligent, species [22]. Because of this kind of smartness, the weaker individuals are not outright losers.
Alternative strategies work not only for mate competition but also apply to food access, social relations, stress response, and other factors. For example while stronger and dominant individuals fight for a patch of food, the submissive ones may wait and watch for opportunities to sneak. This is important to realize because we started with the example of dominant and subordinate males, but it does not imply that the hypothesis being developed applies only to males. We will use the example of alpha male versus subordinate males repeatedly since it is better known to a general reader, has a dramatic appeal, and also much research data are available. But let us take a glance at other examples where being socially smart can partially compensate for not being physically strong. In more intelligent animals higher levels of cognitive abilities are used in tactical deception. A number of interesting observations are depicted by Byrne [23].
The first instance is that of a young male baboon named Paul who used a smart trick to obtain food from a larger and more dominant animal. For example, when an adult female named Mel had just dug up a corm, a prized food item, Paul looked around and seeing nobody looking at them screamed loudly as if someone was harassing him. His mother who was higher ranking than Mel ran into the scene aggressively and chased away Mel, perhaps thinking that Mel was troubling Paul. When his mother was chasing Mel, Paul ate the corm. Paul was observed to use a similar trick more than once but he never used it when in full view of others.
In another instance an adolescent male named Melton played too roughly with an infant and was being chased by the infant’s mother since it screamed. He suddenly gave up running and took an alarm posture staring at a rocky hillside as if he had seen a predator or some other threat. Alarmed by this signal the female gave up the chase and started staring in that direction. Observers confirmed that there was no actual threat present in that direction. Melton had used a tactical deception to escape from a difficult situation.
A primatologist friend of mine, Dr. Anindya Sinha, and his team working on bonnet macaques in India have a number of observations on tactical deception in bonnets, and I have borrowed some stories from their diaries for the readers. These stories belong to a yet unpublished study, and I am waiting eagerly for them to get published.
For example AG, a middle-ranking male, was being allogroomed by a middle-ranking female SU on a tree branch. AG observed TE, another male of a close rank, about to copulate with MU, another female in estrus, about 7 m away on the ground. He intently observed them and gave a very loud alarm call that the troop members usually give to a running pack of dogs. There was not a single dog in sight anywhere. Besides, AG did not scan the area at all but gave the call while looking at TE and MU. The pair became very alert, scanned in all directions, and MU moved off. TE again scanned the area and then warning growled and open-mouth threatened AG, who ignored him completely and resumed his grooming session with SU. Clearly, AG had used a misguiding alarm call to successfully interrupt copulation attempt by a competing male.

In another observation, HS, a young adult male, had recently joined the troop of free-ranging macaques within a zoo that held captive primates as well. HS was consorting with RF, an adult female in estrus. On one occasion, as he approa-ched RF, he suddenly saw PK, the alpha male, approaching them. This was a dangerous situation. HS immediately turned around and gave loud warning growls at a golden langur in a neighboring cage. He then intently observed PK and let RF move ahead, intently watching her but not following her anymore. Here HS had successfully avoided aggression by PK by pretending to be engaged in something else.
Not all acts of deception are successful. At times the deceptive acts appear to be identified as deceptive by the target individuals. This is also a higher-order cognitive task. In another observed story with HS again, he was being chased by JO and NO, two adult females, when he climbed a small tree, looked at a distance, and gave loud alarm calls, normally given to feral dogs. The aggressors, however, ignored the calls completely and continued to direct noncontact aggression towards him. On checking, the observer could not detect any dog or other threat in the vicinity. JO and NO on other occasions responded to threats and alarms normally. But this time they did not as if they knew this was a deceptive alarm call.
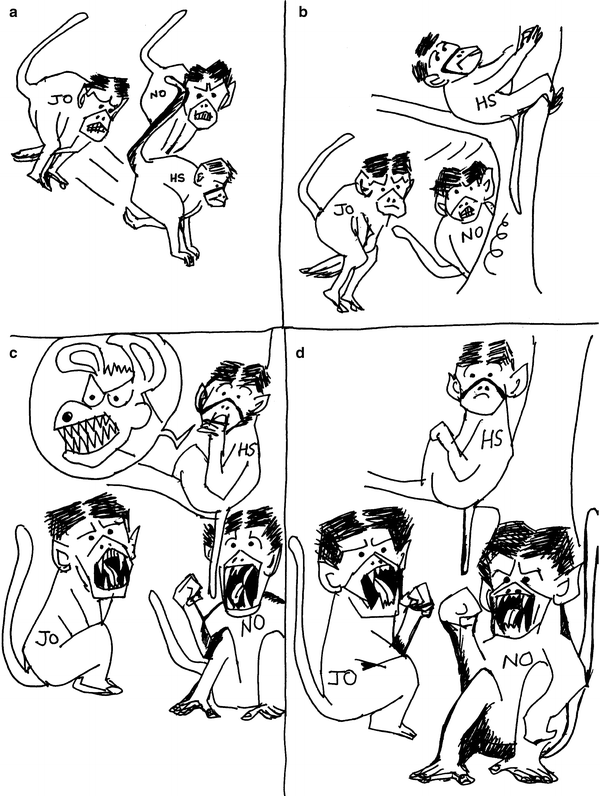
It is interesting to note that in majority of the examples of tactical deception, it was the physically weaker individual that initiated the deceptive behavior. This is not to imply that stronger individuals are not intelligent. It rather reflects the greater pressure on the physically weaker individuals to show social smartness in an attempt to compensate for their weakness. Among the less common examples where a high-ranking individual has initiated tactical deception against a weaker individual, most of the situations are such that strength and dominance alone would have failed to work. For example a dominant individual makes a greeting approach towards a subordinate individual who lets him come close. After being close the dominant individual gives a nasty bite. Here the dominant individual would have been unable to punish the subordinate one if the latter kept on running away. Therefore it is unlikely that dominant individuals are unable to show social smartness. Nevertheless subordinates depend more crucially on deception than the dominant ones. Although examples of weaker individuals using tactical deception or showing social smartness abound, ethologists do not appear to have seriously addressed the question whether there is a negative association or trade-off between physical strength and tactical deception. I suspect such a trade-off, and some of my primatologist friends share the impression, although a rigorous quantitative test of the hypothesis is yet to be done.
In real life whether an individual behaves as a hawk or dove will depend on its social surroundings. We may make a fairly safe assumption that every individual has sufficient behavioral flexibility to choose between the two alternatives. However the relatively stronger individuals will have hawk interactions more frequently than weaker individuals. A consistent difference in these frequencies can make “personalities” or “behavioral syndromes.” A hawk behavior is primarily about aggression, but there are a number of inevitable correlates of physical aggression that are decided by the social structure, foraging, and mating systems of the species. For example aggression, boldness, and exploration are correlated behaviors in fish that form a behavioral syndrome which is shown to have a genetic basis [24]. There can be differential availability of food for dominant and subordinate individuals shaping different metabolic states. A dominant male has greater access to females in a promiscuous species, and therefore, dominant individuals may be sexually more active. Subordinate males, on the other hand, would be sexually more restrained since their overt advances towards receptive females may not be tolerated by dominant males. Aggressive individuals are more injury prone, and therefore, their immune system needs to be more active and ready at the subcutaneous level. Doves, on the other hand, avoid aggression and therefore avoid injuries too which may shape their immune systems differently. In most species submissive display is a very effective way of avoiding actual physical attack. Animal conflicts are highly restrained in terms of inflicting injuries to each other [25]. Therefore although much of the aggression in a group might be directed against the relatively low-ranking individuals, they are much less likely to get actually injured. Injury is more likely when aggression is allowed to escalate, and escalating aggression is typically a hawk characteristic. Doves may partially disinvest from the innate immune and wound healing mechanisms as a parsimonious measure that can be called an immunological “thrift.” If a subordinate individual finds a patch of rich food, there is a risk that a more dominant individual may snatch food from it. However, food once gulped cannot be snatched, and therefore, subordinate animals need to gulp as much as they can before being noticed by a dominant individual. Thus binge eating would be adaptive for a subordinate individual. This implies a peculiar kind of nutritional “thrift” that the physically weak and socially subordinate individuals need to develop. Binge eating directly implies type 1 thrift, but type 2 thrift may also be adaptive for low-ranking individuals who have lower food security. There is a correlated difference in exploring and risk-taking behavior too. The hawks are more likely to be risk takers not only in relation to physical combats but also in relation to foraging. Since physical strength is crucial to their success, they need to obtain better nutrition even if that exposes them to greater predator or any other risk. As a result they are expected to be generally more bold, curious, and exploring their environment with less inhibitions. For weaker individuals, safety is more important than physical strength since physical strength contributes little to their success, but “survive and wait for future opportunities” is a better bet for them. Thus being risk averse in foraging is better for subordinate individuals. Thus the hawk and dove “personalities” go beyond aggression alone and refer to a cluster of correlated behaviors. The two contrasting behavioral syndromes are tabulated in Table 5.1.

Table 5.1
Behavioral syndromes
Hawk/dominant | Dove/subordinate |
|---|---|
Physically stronger | Physically weaker |
Proactive aggression | Aggression avoidance, occasionally reactive aggression |
Snatching | Begging |
Injury prone | Less injury prone |
Optimum eating behavior | Binge eating on opportunities |
Risk takers | Risk and harm avoiders |
Rapid responses to challenge | Relatively slower responses |
Sexually more active | Sexually restrained |
It would therefore be logical to expect that if an individual behaves like a hawk more frequently, its body physiology should be tuned to support a hawk life, and the corollary should be true for dove life too. For example a hawk needs to build and maintain muscle strength. This would need more protein synthesis in muscle for which muscle needs to pick up greater quantities of amino acids as well as glucose. This requires insulin action, and therefore, hawks need to be more insulin sensitive. Physical combat not only requires strength but equally needs agility. Quick reflexes and good nerve muscle coordination are crucial in a fight. Injuries are anticipated in a fight, and the body needs to be prepared for injuries by all possible means. The macrophages should be ready in the subcutaneous tissues to take care of bacteria invading from an open wound. Also the wound healing mechanisms should be more active. This includes the mechanisms of angiogenesis, tissue regeneration, and epidermal growth. For the anticipated loss of blood from injuries, the red cell synthesizing system also needs to be geared up. The social ranks are dynamic, and an alpha position is under continued threat of challenge by some other individual sooner or later. This has two important implications. One is that the stronger individuals need to be more of risk takers to gain and retain an alpha position since the possible rewards from risky behaviors are very high. The other implication is that the hawks need to be sexually more active since they need to reproduce as much as they can before their dominant status is threatened. Not surprisingly sex and aggression mechanisms have a substantial mechanistic overlap. Some of the sex hormones such as testosterone in males have proaggression effects, whereas the effects of estradiol on female aggression are complex and mixed [26–29]. The set of neurons in the hypothalamus involved in attack and in mating also overlap [30]. The link between aggression and reproductive capacities is perhaps much wider. It is recorded in colonial wasps that a new queen cannot have sufficient fecundity unless it displays substantial dominance aggression [31].
The physiological requirements of the dove or subordinate animals are diametrically opposite. Since they are weaker, aggression is counterproductive for them, and they need to completely suppress proactive aggression and as far as possible avoid circumstances necessitating reactive aggression. As a result they become less dependent on strength and more on social manipulation. They can afford to disinvest from muscle building. Amino acid and glucose uptake of muscle can be reduced as a parsimonious or thrifty strategy. But simultaneously cognitive functions of the brain need to be enhanced. Since unlike most other tissues that can use fatty acids for energy, the brain is more specifically dependent on glucose; it is more critical to assure glucose supply to the brain, whereas glucose supply to the muscle may be compromised. Doves need to be more socially supported, and they indulge in activities such as allogrooming that help making alliances and ensuring greater social support. Unlike the dominant individuals, they are expected to be more sexually restrained with lower levels of sex hormones. Since they can effectively avoid fights by increasing submissive displays [25], they are less injury prone and can partially disinvest from peripheral innate immunity, wound healing mechanisms, angiogenesis, and erythropoiesis as compared to a dominant individual. They also need to be more risk avert. Social manipulation is a relatively slow process as compared to a physical fight where split-second decisions are crucial. As a result the responses to challenges by doves could be more sluggish than hawks.
Do our expectations match with data? If our conjunctures are true, a number of subtle physiological differences should be observable between hawks and doves in animal societies. Ethologists generally do not classify individuals as hawks and doves, but in social species where a higher social rank is mainly achieved by physical strength and aggression (notably this is not true for all species), the dominant individuals can be taken equivalent to hawks and subordinate as doves. Our classification should depend on “effective” aggression. Often an alpha male does not have to behave overtly aggressively with other members of the troop very frequently since his dominance is already accepted by others. An intermediate individual may show more frequent aggression which may or may not be too effective in raising its status. It is therefore better to use social ranking that reflects effectiveness of aggression rather than using the frequency of aggression to identify hawks and doves. A note of caution is that this cannot be generalized across species. In primates, for example, physical aggression has a variable role in attaining and maintaining social dominance. The role of aggression can also differ between sexes in a given species. In olive baboon males, overt aggression is the key to attaining dominance, and threat and intimidation are necessary to maintain it [20, 32]. On the other hand, among females of common marmosets, aggression is rare, and dominance does not appear to be dependent on it [32, 33]. Nevertheless both aggression and dominance have important roles in determining reproductive success, although their relative importance and correlation with each other may vary across genders and species. Since our focus is on effective aggression and we are using social dominance as a marker, the discussion below refers to species in which aggression is the predominant determinant of dominance.
In recent years there have been an increasing number of studies on the endocrine and metabolic states of primates and other animals in the wild or socially reared captive colonies. Animal models of insulin resistance are not new. Even primate models for obesity and insulin resistance are known for a long time. In the earlier models some or the other obesifying interventions were deliberately used to induce obesity and insulin resistance. These include dietary manipulations, restricting physical activity, or specific brain lesions interfering with energy homeostasis. Animals do develop obesity and insulin resistance in response to such interventions. However, it is known now that such interventions are not necessary prerequisites of insulin resistance. A significant fraction of individuals in a social group of primates is shown to become insulin resistant around puberty even in the absence of any such interventions [34]. This was demonstrated in bonnet macaques that had normal diet, physical activity, and social interactions. Insulin resistance that developed among the 15% of individuals by peripubertal age was not contributed by high-calorie intake, physical inactivity, or obesity. Interestingly the proportion of insulin-resistant individuals at the physically active peripubertal age was not different from that in the significantly older and sedentary age class [34]. It is possible therefore that insulin resistance in a proportion of individuals is an inherent part of the complex social life in primates and may not have a pathological origin at all.
Data coming from wild or captive groups of animals are fragmentary. Some studies have data on diet but not social interactions, and some others that have looked at social interactions have not reported morphometry and so on. Among the physiological parameters, some studies have monitored corticosteroid levels but not insulin. Some focus on cholesterol but do not estimate plasma glucose. It is possible, nevertheless, to join the pieces together and try to make a coherent picture. Generalizing over a wide variety of species of vertebrates, we can detect certain common patterns. The dominant and aggressive individuals of both sexes typically have high levels of sex hormones, low serotonin and higher dopamine activity in the brain, lower plasma cholesterol, and corticosteroids as well as low plasma insulin. The subordinate ones have the opposite picture of low sex hormones, high serotonin, low dopamine, and higher plasma levels of corticosteroids and cholesterol, more specifically low HDL and high LDL cholesterol [32, 35–57]. There are exceptions though, such as subordinates having lower corticosteroid levels than dominant individuals, but they come from species in which aggression is less important in determining dominance [32]. Fewer studies have looked at the relationship between dominance hierarchy and insulin. One study on wild chimpanzees in the Kibale National Park showed that subordinate individuals have higher urinary C-peptide [58] which reflects higher insulin production and thereby insulin resistance. This pattern was consistent over 13 of 14 seasons monitored and independent of high or low fruit availability. This appears to have come as a surprise to these researchers. They expected the urinary C-peptide to reflect calorie intake and therefore expected the high-ranking individuals to have higher levels. The expected association of calorie intake to insulin levels was seen across seasons. In seasons of better availability of fruits, all individuals had higher levels of C-peptide. This is consistent with other studies showing higher obesity and insulin levels with increased availability of food [59]. But across the society, the effect of social ranking was stronger than the effect of calorie intake. This might be extremely important and relevant to understanding insulin resistance syndrome in humans as we will discuss in the next chapter. Another study looked at insulin-like growth factor 1 (IGF-1), which plays important roles in protein metabolism, muscle physiology, wound healing, erythropoiesis, and immunity and found that social subordination is associated with suppressed levels of IGF-1. This finding in baboons is also relevant since IGF-1 has an important role in controlling insulin resistance as we will see in a later chapter. This study also asserts that the suppressed IGF-1 response is consequential rather than causal to subordination [49]. It is important to realize that some of these studies are on wild primate groups, indicating that these physiological effects are not an artifact generated by captive conditions. The trend in the physiological gradient along the social hierarchy is similar in captive and wild primates. In birds, individuals with a more timid personality have been shown to have greater oxidative stress [60]. Subordinate animals also have higher resting blood pressure [37, 61, 62]. Thus in comparison with hawks, the endocrine and metabolic picture of the doves goes close to metabolic syndrome in humans. In the wild it is rare for subordinate individuals to become overt diabetic and develop clinical complications typical of diabetes. The magnitude of the endocrine and metabolic changes in animals is generally not large enough to cause the pathophysiology commonly seen in the human metabolic syndrome, but the direction of the change is the same. Interestingly higher levels of cholesterol, corticosteroids, and insulin along with moderate insulin resistance are recorded in subordinate individuals in spite of having lower caloric intake as compared to dominant individuals [37, 58]. This apparently counterintuitive example is important in the context of metabolic syndrome in humans as we will see later. Further our expectation that physiological response to social challenges of a hawk would be faster than a dove is also supported by evidence. The stress responses of dominant individuals are typically rapid and sharp. Those of subordinates are sluggish, although they have higher basal levels of stress hormones [37].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


