6 Endocrinology
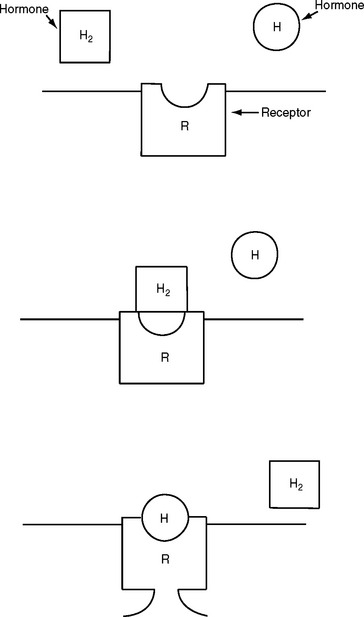
Hormones are chemical substances secreted from glands through ducts and sent to other tissues through the bloodstream or secreted by glands with no ducts into the surrounding tissue. These chemicals affect only certain tissues and therefore evoke a specific response. Tissues cannot respond to a hormone unless that tissue contains receptors for that hormone. Receptors act like gates on the surface of the cell, with the hormone acting like a key; if the key fits the gate, a response occurs within that cell (Figure 6-1).
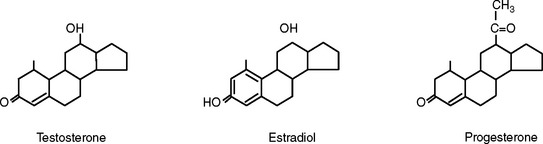
The other class of hormones is steroids. The steroid hormones—testosterone, estrogen, progesterone, and cortisol—all have a very similar chemical structure (Figure 6-2). Steroid hormones do not differ between species, so they can be used interchangeably for treatment and measured in the blood of any species using any species’ assay.
An important concept in the control of hormone secretion is feedback. Increasing concentrations of a given hormone in the bloodstream will usually “feed back” to the stimulus that caused release of the hormone, thereby stopping further production. An example is testosterone secretion. The release of interstitial cell–stimulating hormone from the pituitary gland stimulates the release of testosterone from the testes. Increasing concentrations of testosterone in the bloodstream signal the pituitary to stop release of interstitial cell–stimulating hormone, which in turns stops further production of testosterone. This is called a negative feedback loop. When concentrations of testosterone in serum fall, the pituitary is again stimulated to release interstitial cell–stimulating hormone and the cycle starts again. This is an important concept because it alters how we use hormones for therapy. Any hormone introduced into the body may exert negative or positive feedback on glands and tissues, altering the animal’s ability to produce or secrete that hormone now and in the future. Caution must be employed when introducing hormones into the body.
I. ENDOCRINE TISSUES AND THEIR PRODUCTS
A. HYPOTHALAMUS
The hypothalamus lies at the base of the brain (Figure 6-3). It has a unique blood system connecting it to the pituitary gland, which lies beneath it. This blood system, the hypophysial portal system, carries hormones from the hypothalamus directly to the pituitary without circulating it elsewhere in the body. This means that only very tiny concentrations of hypothalamic hormones need be produced to stimulate an effect in the pituitary gland.
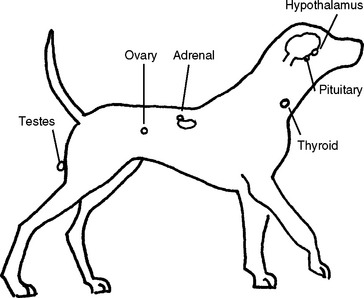
All hormones produced and released from the hypothalamus cause a direct positive or negative effect on the secretion of hormones from the pituitary. The hypothalamic hormone of greatest interest to small animal reproduction is gonadotropin-releasing hormone (GnRH), a very small protein hormone. The stimuli that cause release of GnRH are not well defined but probably include environmental stimuli and feedback from secretion of pituitary hormones. GnRH causes release of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) from the pituitary. GnRH is used clinically to induce ovulation (release of eggs from the ovary), and related compounds have been described for inducing estrus in bitches and, with long-term therapy, for suppressing estrus. It has been described as a reversible contraceptive in males in some species. GnRH also can be used in males to cause release of LH, which in turn will cause release of testosterone. This is useful if there is a need to measure testosterone production in a dog and can be used occasionally to increase libido in male dogs.
B. PITUITARY
The hormones of the posterior lobe are oxytocin and vasopressin. Vasopressin is involved in kidney function and will not be discussed here. Oxytocin is a protein hormone. It is released secondary to neurologic stimulation. The two stimuli best documented to cause oxytocin release are pressure of the head of a puppy into the cervix during whelping (Ferguson’s reflex) and suckling of the mammary glands by the pups. In both instances, nerve cells are stimulated to fire, including those in the hypothalamus that produce oxytocin. Stimulation of these nerves causes production of oxytocin and release of the hormone from its site of storage in the posterior pituitary. Oxytocin causes synchronized contractions of the uterus during parturition and contributes to further dilation of the cervix. Oxytocin receptors are not always present on the uterus. Stimulation of the uterus with another hormone, estrogen, increases the number of oxytocin receptors. For this reason, oxytocin is not used in some disease conditions that require contractions of the uterus to empty it of discharge. Oxytocin can cause very strong uterine contractions if given at the right time and should always be used with care and only on the advice of a veterinarian. Clinical use of oxytocin during whelping is discussed in Chapter 12.
During lactation, oxytocin stimulates letdown of milk into the mammary glands. Oxytocin does not stimulate milk production, only milk letdown. Lack of milk production (agalactia) is uncommon in dogs (see Chapter 18). If oxytocin is going to induce milk letdown, it will work within seconds of intranasal or systemic (injected) administration.
FSH is a large protein hormone secreted both in males and females. It is named for its role in the female, in which it stimulates maturation of ovarian follicles in preparation for ovulation (see Chapter 7). In males, FSH stimulates production of a protein in the testes that binds testosterone tightly within the testis to promote spermatogenesis. FSH is not available as a commercial product and has no clinical use at this time.
Another large protein, LH, also is named for what it does in the female. Luteinization in dogs occurs in the ovarian follicles around the time of ovulation (see Chapter 9). LH could be given to dogs to induce ovulation but is not commercially available. A human hormone produced during pregnancy, human chorionic gonadotropin (hCG), when administered to dogs, binds to LH receptors and can be used to induce ovulation. LH is also called interstitial cell stimulating hormone, named for its function in males. In male dogs, LH stimulates release of testosterone from the testes, which is necessary for continuing production of spermatozoa. LH production is controlled by a feedback loop with testosterone.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


