CHAPTER 57 Effects of Nutrition on Reproduction in Dairy Cattle
Walton, as cited by Coppock,1 predicted that individual cow yields of more than 31,000 kg of milk per year were attainable, with the potential for herd averages of 16,000 kg of milk per year. Breed averages for milk yield increased 40% to 60% for first-lactation cows between 1960 and 1988.2 With continued genetic progress and improvements in management systems, Walton’s predicted levels of milk production are reasonable and achievable goals.
As milk production increases, fertility of lactating dairy cattle, as a general rule, decreases. A survey of commercial herds in North Carolina showed a decrease in conception rate from 52% to 32% in a comparison of the lowest-producing (n = 425) and the highest-producing (n = 53) herds.3 Days open and days to first service were similar for both groups. More recently, Washburn and associates4 summarized Dairy Herd Improvement Association (DHIA) data from 10 states in the southeastern United States. In the period 1976 to 1999, annual milk yield per cow increased 1800 kg, while days open increased from 112 to 152 during the same time period. The herds also experienced an increase in services per conception (from a mean of 1.9 to 2.9). Lucy5 completed an extensive review and suggested that high levels of milk production are not the sole cause of reduced reproductive performance. Indeed, overall management including housing, health, and nutrition practices also affects fertility of lactating dairy cattle. These findings emphasize the importance of management in maintaining reproductive efficiency as herds achieve higher levels of milk production.
Nutritional status and nutritional management are essential components of overall herd management. The nutritional status of the herd also affects the reproductive soundness and efficiency of the herd. Bauman and Currie6 described the hierarchy of nutrient utilization by the lactating dairy cow. Within this ranking, reproduction lists below maintenance (necessary for survival), production (milk produced for the young), and growth. Royal and coworkers7 summarized strategies to improve fertility in dairy cattle, including genetic, endocrine, and nutritional approaches. Ferguson8 categorized nutritionally associated reproductive problems as primary or secondary. Primary conditions of malnutrition affecting reproduction include clinical deficiency and excessive intake of a particular nutrient. Secondary conditions comprise those in which other constraints result in a nutritional imbalance. Therefore, a suboptimal plane of nutrition can result in impaired reproductive performance. This observation holds true for both the developing replacement dairy heifer and the lactating dairy cow.
PUBERTY
Gill and Allaire9 analyzed DHIA records from Ohio and concluded that the optimal age for Holstein heifers at first calving for total lifetime production was between 23 and 24 months. More recent research demonstrated that heifers calving early (22.9 months of age) produced more milk than those calving at an older age (26.5 months).10 To achieve an average age at first calving of 24 months, heifers must reach puberty by 8 to 9 months of age. This timing would allow heifers to establish cyclicity before the breeding period. Establishing cyclicity before breeding has been shown to improve fertility. For example, the conception rate for heifers at pubertal estrus was 21% lower than at their third estrus.11 An excellent plane of nutrition is critical for heifers to reach puberty and first calving at 9 months and 24 months of age, respectively. Lammers and colleagues reported that heifers needed to gain 820 g/day (average daily gain from birth to calving) to achieve body weight and age targets at first calving.12
A number of factors that include body weight, body size, plane of nutrition, breed, and season influence age at onset of puberty. Social environment and phase of the moon also have been reported to alter onset of puberty.13 Nutrition, because it influences body weight and size, contributes to the onset of puberty. The age at which a heifer begins regular estrous cycles also is correlated with the rate of gain from birth to puberty. Bortone and colleagues14 reported that heifers fed 115% of National Research Council (NRC) recommendations from 3 to 12 months of age were 22 days younger at the onset of puberty than control animals fed 100% of NRC levels. The correlation between gains in body weight and age at puberty indicates that an increased growth rate in heifers reduces the age at puberty.15,16 The study of Bortone and colleagues14 also found that the body weight of heifers at the onset of puberty was the same for faster-growing heifers as for controls. These data indicate that body weight can be used to predict the onset of puberty and establish rate of gain.
Feeding heifers to reach target weight at a selected age for a given genotype is a practical management tool to ensure high potential fertility. Nutrient intake that supports growth and development is essential to achieve a target weight by 9 months of age. Nutrient intake is controlled by growth rate, fiber and energy concentration (or density) of the diet, and the dry matter content of the diet. Tomlinson and associates,17 using prepubertal and pubertal heifers, found maximal dry matter intake at a neutral detergent fiber level of 41% and an acid detergent fiber level of 20% of the dietary dry matter. Quigley and coworkers18 reported that intake could be limited by diets containing greater than 23% acid detergent fiber. Intake with high-fiber diets may be limited by the physical constrictions of rumen fill and distention. A decreased rate of fermentation and rate of passage of high-fiber diets also can limit dry matter intake.19 Nutrient density and fiber content of the diet should be evaluated when animals do not reach the production goals of puberty, breeding, and first calving.
Feed additives also have been used in the diet of developing heifers. Sodium bicarbonate and yeast have been studied for use in the diet of young calves. Observed effects on intake, ruminal fermentation, and health have been variable and nonconclusive.20 The use of ionophores generally improves growth rates, as well as feed efficiency, of ruminants. Two compounds commonly used are lasalocid and monensin. Feeding ionophores has been shown to decrease age at first breeding and calving, primarily because the growth rate to adequate body size and weight is accelerated. It has been hypothesized that monensin decreases age at puberty by a mechanism independent of weight gain.21 Estradiol implants also have been found to improve feed efficiency in prepubertal Holstein heifers.11
Plane of nutrition also can influence the onset of puberty. Plane of nutrition can inhibit the onset of puberty in heifers at an adequate body weight, size, and age. Gonzalez-Padilla and associates22 demonstrated that 14-month-old heifers could be maintained in a prepubertal state and then suddenly made to ovulate by increasing nutrient intake. It generally is accepted that the physiologic cause of the delay is the result of inhibition in pulsatile release of gonadotropin-releasing hormone (GnRH) from the hypothalamus and the subsequent release of luteinizing hormone (LH) from the pituitary gland. It also has been demonstrated that GnRH secretion is extremely sensitive to the nutritional status of the animal. Furthermore, insulin may play a role for energy-yielding nutrients, as it binds receptors in the region of the brain thought to regulate GnRH secretion.23 Conditions such as disease and parasitism can influence nutritional status by altering intake or absorption of nutrients. Bumgarner and coworkers24 reported that calves treated with anthelmintics in both mid-June and mid-July had a reduction in fecal oocyte counts and increased daily gain. Numerous other studies demonstrated improved weight gains from the use of anthelmintics.25 Respiratory disease that results in severe lung damage can also result in poor performance of growing dairy heifers. A sudden onset of disease can result in a transient period of anestrus.
LACTATING COW
As discussed earlier, Bauman and Currie6 described the need of lactating cows to prioritize nutrient utilization. In general, energy is the nutrient of primary concern relative to supporting maintenance, lactation, and reproduction. Imbalances or deficiencies of protein, minerals, or vitamins also have been reported to adversely affect the reproductive function of lactating dairy cows. Nutritional imbalances or deficiencies are most important during the period immediately preceding parturition through conception. This section focuses on the influence of nutritional status at critical points throughout lactation.
The nutritional balance of a dairy cow during the periparturient period can have a tremendous influence on fertility and reproductive efficiency. During the final 30 days of gestation, lactation is initiated, with production of colostrum and final growth of the fetus. At day 270 of gestation, the uterus and fetus require greater than 1600 kcal per day.26 Concurrent with increased nutrient requirements, the prepartal cow experiences a marked decrease in dry matter intake. Severe anorexia or an imbalance of nutrient intake can predispose the animal to a number of metabolic diseases that constitute the parturition disease complex.27 These diseases are not independent but can increase the risk of subsequent problems. For example, Curtis and coworkers28 reported that cows with hypocalcemia were at increased risk of suffering dystocia, retained fetal membranes, and ketosis. Dystocia and retained fetal membranes are predisposing factors for postparturient uterine disease. Markusfeld27 documented that cows experiencing postparturient uterine disease had a decreased milk yield for the lactation and increased number of days open. Retained fetal membranes also have been influenced by selenium and vitamin E supplementation. Cows that experienced retained fetal membranes may have had a lower plane of nutrition. Disrupted intake accompanied by disrupted energy metabolism can result in an increased level of circulating ketone bodies. Miettinen29 reported that cows with higher concentrations of circulating ketone bodies had more days to first service, decreased first service conception rates, and more days open. More recently, cows with serum beta-hydroxyl butyrate concentrations of 1100 μmol/L in weeks 1 and 2 of lactation were found to have increased risk of conception failure.30 These studies make a significant statement regarding the importance of periparturient health and metabolism on fertility and reproductive efficiency during subsequent lactation. The primary modulator of reproductive function appears to be energy intake and the energy status of the animal.
Energy
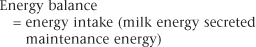
With initiation of lactation, the acceleration in milk production exceeds the increases in dry matter intake. Kertz and colleagues31 showed that dry matter intake increased gradually, to peak at week 10 after calving. Lactating cows consume inadequate dietary energy to meet peak milk production. As a result, early-lactation cows experience a negative energy balance (Fig. 57-1). On the basis of energy calculations, cows in early lactation can experience as much as a 20-Mcal deficit of net energy. Cows compensate during this delay in energy intake by mobilizing stored body energy. For the early postpartum cow, this energy is generated primarily from stored adipose tissue. One kilogram of body fat contains 6 Mcal of stored energy.31 Therefore, early-postpartum dairy cows must rely on oxidation of body fat for energy to compensate for inadequate energy intake.
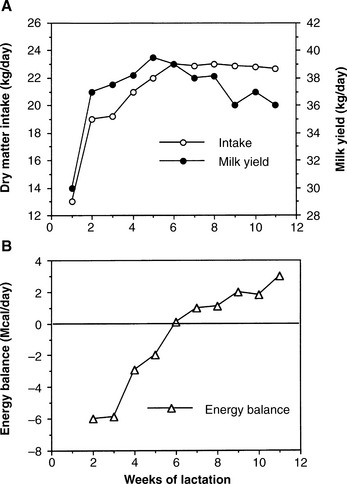
The nutritional challenge of negative energy balance is significant and can adversely affect the reproductive performance of the dairy cow (Fig. 57-2). A similar effect is recognized in most species. This response by the animal may represent a mechanism to conserve energy during periods of starvation. This same mechanism, however, allows the animal to synchronize reproductive events with periods of positive nutritional status. The timing of these nutritional challenges is an important consideration in management, in view of the need to have high fertility and conception rate during early lactation. The inhibitory factors that prevent pregnancy are only partially understood. Negative energy balance can increase the period of postpartum anestrus and decrease fertility at first and subsequent inseminations, depending on the severity of the imbalance.
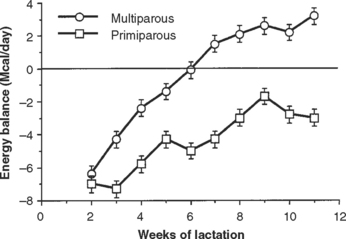
Fig. 57-2 Average daily energy balance for multiparous (n = 35) and primiparous (n = 15) Holstein cows.
(From Lucy MC, Staples CR, Thatcher WW, et al: Influence of diet composition, dry-matter intake, milk production and energy balance on time of postpartum ovulation and fertility in dairy cows. Anim Prod 1992;54:323.)
Cows should experience the first postpartum ovulation by 30 days after calving. Cows are not inseminated at this time because of incomplete involution of the uterine tissues. It is important, however, to minimize the length of postpartum anestrus for two reasons. First, cows with long intervals to first ovulation will be anestrous during the breeding period. Conception will therefore not occur because the cow does not express estrus and does not ovulate. Second, a shorter time to estrus will enable the cow to have multiple ovulations before the first insemination. The estrous cycle has a cleansing effect on the uterus, with increased uterine contractility and relaxation of the cervix. Therefore, as shown in a large study conducted in Florida,32 cows with prebreeding estrus had higher first-service conception rates (47%) compared with cows with no prebreeding estrus (34%).
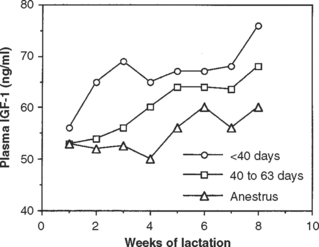
To achieve the maximum number of estrous cycles before breeding, cows must ovulate during the early postpartum period (<21 days in milk). Research has shown that cows will ovulate approximately 10 days after reaching the lowest point of negative energy balance, the nadir.33 Growth of follicles and ovulation are dependent on the pulsatile secretion of LH.34 In cattle in severe negative energy balance, the secretion of LH is inhibited.35 Disrupted or decreased LH secretion slows the growth and development of the follicle, which delays ovulation.36 Insulin and insulin-like growth factor-I (IGF-I) also are required for normal follicular growth and ovulation. Cows in negative energy balance have reduced levels of IGF-I. This is an important relationship, given that IGF-I amplifies the effects of LH on the ovary through potentiating the signaling mechanism for LH.37 Therefore, the actions of LH are decreased in cows in negative energy balance because lower IGF-I leads to reduced effectiveness of LH. Consequently, follicles in cows with extremely low IGF-I do not develop normally, and ovulation is delayed because LH is less active. This effect was demonstrated in a study by Thatcher and coworkers38 (Fig. 57-3). Cows that had first ovulation before 40 days post partum had the highest concentrations of plasma IGF-I.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


