Fig. 2.1
Cuff placement for the femoral artery
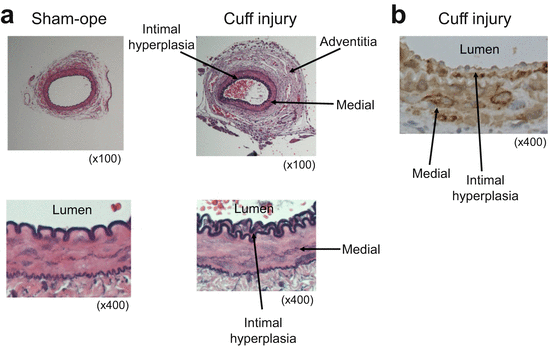
Fig. 2.2
Representative arteries after sham-operated or cuff placement (a) and alpha smooth muscle actin staining for cuff-injured artery in C57B6 mice (b)
2.5 Etiology of Cuff-Induced Neointimal Formation
Since cuff-induced neointimal formation showed positive staining with an α-SMC antibody (Fig. 2.2b), the neointima appears to be almost entirely composed of SMCs. Neointimal formation induced by cuff placement can be considered as a consequence of migration and/or proliferation of the SMCs induced by some factors such as PDGF-BB or inflammatory cytokines (Fig. 2.3). Although the exact mechanisms underlying the neointimal formation in cuff-induced injury models are still uncertain, I would like to refer to reports from the literature of studies carried out using genetically manipulated mouse models to discuss the mechanisms of neointimal formation (Table 2.1).
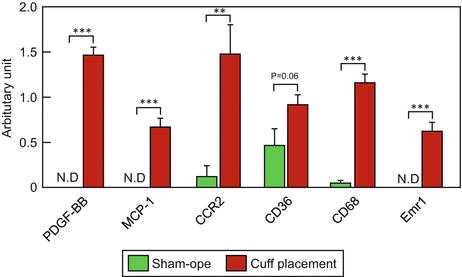
Fig. 2.3
Comparison of the gene profiles between sham-operated artery and cuff-injured artery
Table 2.1
Representative cuff placement in genetically manipulated mouse models
Gene | Location | Neointimal formation | References | |
|---|---|---|---|---|
LR11-KO | Carotid artery | Reduced | Arterioscler Thromb Vasc Biol | 2007 [38] |
Rag-1-KO | Carotid artery | Increased | Arterioscler Thromb Vasc Biol | 2002 [45] |
CD4-KO | Carotid artery | Reduced | J Am Heart Assoc | 2013 [50] |
eNOS-KO | Femoral artery | Increased | J Clin Invest | 1998 [34] |
iNOS-KO | Carotid artery | Reduced | Circ Res | 1999 [59] |
sEH/ApoE double-KO | Femoral | Reduced | Arterioscler Thromb Vasc Biol | 2010 [61] |
sEH/ApoE double-KO | ||||
Tlr4-KO | Femoral | Reduced | Circulation | 2002 [64] |
Tlr2-KO | Femoral | Reduced | Cardiovasc Res | 2005 [65] |
CatK-KO | Carotid | Reduced | Hypertension | 2014 [66] |
AT1 receptor-KO | Femoral | Reduced | Circulation | 2002 [74] |
AT2 receptor-KO | Femoral | Increased | Circulation | 2002 [74] |
Mas-KO | Femoral | Increased | Hypertension | 2014 [78] |
Cu/ZnSOD-KO | Femoral | Unchanged | Journal of atherosclerosis and thrombosis | 2011 [81] |
HAS2Tg (SMC-specific) | Femoral | Increased | PLoS One | 2013 [39] |
IRS1-KO | Femoral | Increased | Circulation | 2003 [85] |
IRS2-KO | Femoral | Increased | Circulation | 2003 [85] |
2.5.1 Factors Directly Inducing SMC Proliferation and Migration
Cuff replacement around the femoral artery induced neointimal hyperplasia, which was exclusively composed of SMCs, as mentioned above. Platelet-derived growth factor (PDGF)-BB, which is mainly secreted from platelets, is one of the most potent factors inducing migration and proliferation of the SMCs after balloon angioplasty. The expression levels of PDGF-BB mRNA in the cuffed artery were significantly increased as compared to those in the sham-operated artery (Fig. 2.3). An antagonist of the PDGF-BB receptor was shown to suppress the proliferation of vascular SMCs after balloon-induced injury in a rat model [36]. PDGF-BB induces vascular SMC proliferation via, at least in part, regulation of the cell cycle. Schwaiberger et al. demonstrated that local treatment with a cyclin-dependent kinase inhibitor inhibited cuff-induced neointimal formation, accompanied by reduction of the phosphorylation level of signal transducer and activator of transcription 3 (STAT3), but not of Akt, ERK1/2, or p38MAPK activation [35]. These data suggest that PDGF-BB-STAT3 signaling is involved in cuff-induced neointimal formation.
In addition to PDGF-BB-STAT3 signaling, PDGF-BB has been reported to augment the migration of SMCs through the LR11, a member of the LDL receptor family/urokinase-type plasminogen activator receptor (uPAR) pathway [37]. Ohwaki et al. found that balloon-injured intimal SMCs showed strong expression of LR11 in rat arteries. High-fat diet-fed LR11-KO mice showed a decrease of cuff-induced neointimal formation [38]. In in vitro experiments, the secreted soluble form of LR11 (soILR11) promoted the migration of THP-1 induced by PDGF-BB, an effect that was completely canceled by the anti-uPAR antibody [38]. These data suggest that PDGF-BB induced by cuff-induced injury is one of strongest regulators of neointimal formation.
The ECM has been reported to be a key player in arterial remodeling after vascular injury [4]. Hyaluronan 2 (HAS2), which is a primary component of the ECM, is expressed in neointimal lesions in humans with atherosclerosis and at sites of wire-induced injury of the arteries in mice. Mice overexpressing the murine HAS2 gene specifically in the vascular SMCs (cHAS2/CreSM22a mice) showed markedly enhanced cuff-induced neointimal formation, with augmentation of SMC migration and proliferation, and production of inflammatory cytokines and ROS [39]. Consistent with these data, in a wire-induced injury model, a HA synthesis inhibitor markedly suppressed neointimal formation. The ECM induced by arterial cuff-induced injury promotes migration of SMCs into the intima, leading to neointimal formation.
2.5.2 Roles of BM-Derived Cells in Cuff-Induced Neointimal Formation
The roles of bone marrow (BM)-derived cells in the pathogenesis of atherosclerosis have been extensively studied [40]. Fluorescence in situ hybridization (FISH) analysis has revealed that female-derived BM cells transplanted into male mice were detected in the neointima of male mice (Fig. 2.4). Following BMT from green fluorescent protein (GFP)-transgenic mice to ApoE-KO mice, GFP-positive cells were confirmed in the vascular neointima (4–10 %) and media (5–32 %) in the latter mice. Following bone marrow transplantation (BMT) from LacZ mice to wild-type (WT) mice, a number of LacZ-positive macrophages were found in the neointima (25.7 %), media (7.3 %), and adventitia (73.7 %) of the arteries of the WT mice at 4 weeks after cuff placement [33]. It was worthy of note that more marked infiltration of the adventitia by macrophages was observed after injury induced by cuff placement than after wire injury or ligation injury.
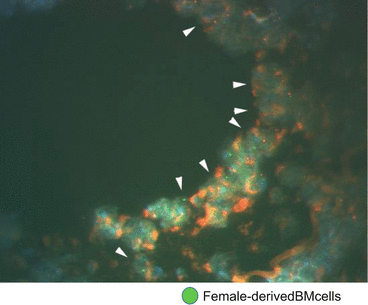
Fig. 2.4
Fluorescence in situ hybridization (FISH) of cuff-injured artery in male mice transplanted female-derived BM cells
Xu et al. transplanted the BM of GFP-transgenic mice into LDLR-KO mice to identify the cell lineage in the lesion [41]. Two weeks after cuff placement following a high-fat diet for 4 weeks, atherosclerotic lesions developing in the intima predominantly consisted of a massive accumulation of foam cells with a number of α-SMA- and GFP-positive cells. In addition to macrophages, adventitial small vessels also showed positive staining for both the endothelium-specific marker CD31 and GFP in mice transplanted with BM obtained from GFP-transgenic mice [41]. Most of the macrophages are GFP-positive, and some of the SMCs and ECs are also GFP-positive. In a cuff-induced vascular injury model, BM-derived cells are recruited to the adventitia and differentiate into macrophages, SMCs, and endothelial cells. Treatment with a blocker of M-CSF, which guides differentiation into macrophages, caused a prominent decrease of macrophages, and inhibition of the PDGF receptor suppressed the recruitment of SMCs to the adventitia after cuff placement [42]. These data suggest that the adventitia plays a pivotal role in neointimal formation induced by cuff placement. Scott et al. demonstrated that the adventitia is important in the first wave of growth after angioplasty following occurrence of neointimal hyperplasia [43].
2.5.3 Roles of T Cells and B Cells in Cuff-Induced Neointimal Formation
The importance of T and B lymphocytes in the process of restenosis after PCI has been pointed out [44]. Rag-1-KO mice, which lack mature B and T cells, have been demonstrated to show increased thickness of cuff-induced neointima in the carotid artery. Reconstitution of the Rag-1-KO mice with B cells from WT mice reduced the neointimal formation. Moreover, both IgG and IgM were detected in the cuff-injured carotid arteries of reconstituted Rag-1-KO mice with B cells [45]. These data suggest that an increase of immunoglobulin by activation of B cells exerts a protective action against intimal thickening. The neointimal area and intima/media ratio were significantly reduced in mice treated with immunoglobulin administered intraperitoneally for 5 consecutive days starting 1 day prior to cuff placement. On the other hand, immunoglobulin could not suppress cuff-induced neointimal formation, when the treatment was commenced 3 days after cuff placement [46]. When pooled mouse IgG or IgM was given to Rag-1-KO mice by intravenous injection, a significant reduction of intimal thickening was observed as compared with that in the untreated Rag-1-KO mice. Immunoglobulin treatments modify the serum complement C3 profile, and the amount of complement C3 was decreased in the injured arteries. Depletion of complement C3 in the Rag-1-KO mice significantly decreased the degree of intimal thickening [47]. These data suggest that IgG, IgM, and complement C3 are involved in the modulation of the neointimal hyperplasia response to cuff-induced injury.
Moreover, treatment with immunoglobulin significantly enhanced the secretion of interleukin (IL)-10, suggesting activation of T cells by immunoglobulin [46]. Rag-1-KO mice reconstituted with T cells from WT mice showed a reduction of neointimal formation after cuff placement [48]. Dimayuga et al. carried out a detailed examination of the T-cell fraction. Splenic CD8+CD25+ T cells and CD8+CD28+ T cells, but not CD4+CD25+ and CD4+CD28+ T cells, were also significantly increased after arterial injury in the WT mice. Rag-1-KO mice given CD8+ T cells showed a significant decrease of neointimal formation as compared to Rag-1-KO mice not given the cells. On the other hand, transfer of CD4+ T cells was not associated with inhibition of the neointimal formation [49]. Neointimal formation induced by cuff placement was significantly reduced in CD4-KO mice as compared with that in the WT mice, because of the higher percentage of CD8+ T cells. Moreover, adoptive transfer of CD8+CD28hi T cells into recipient Rag-1-KO mice significantly reduced neointimal formation as compared to that of CD8+CD28+ T cells [50]. Although both CD8+ T cells and CD4+ T cells are activated in response to arterial injury, CD8+ T cells, which constitute at least a fraction of the CD8+CD28hi, are mainly involved in the inhibition of cuff-induced neointimal formation.
2.5.4 Roles of eNOS and iNOS in Cuff-Induced Neointimal Formation
Nitric oxide (NO) is an important vascular regulatory factor that is generated by the enzyme nitric oxide synthase (NOS) [51]. Three different isoforms of NOS are recognized: among these is endothelial NOS (eNOS), which is constitutively expressed mainly in the vascular endothelial cells [52]. eNOS can be an important factor modulating vascular endothelial function and is activated by acetylcholine (ACh) and insulin. eNOS-KO mice exhibit impaired ACh-induced vascular relaxation [53]. The second isoform is inducible NOS (iNOS), which cannot be detected in normal tissue, but is expressed in several cell types, including macrophages and vascular SMCs, after cytokine stimulation [54], and the third isoform is neuronal NOS (nNOS), which is constitutively expressed mainly in the nervous tissues and skeletal muscle type II.
eNOS-KO mice exhibit increased neointimal formation following cuff placement [34]. Consistent with these data, the neointimal formation induced by ligation is also significantly more pronounced in the eNOS-KO mice than in the WT mice [55]. Since antiplatelet and antihypertensive treatments cannot attenuate the progression of neointimal formation, the neointimal hyperplasia observed in eNOS-KO mice is produced by the direct action of eNOS and not mediated by thrombus formation or high blood pressure [33]. When adenovirus-mediated human endothelial constitutive NOS cDNA (AdCMVceNOS) was transduced into the rat carotid artery after balloon injury, the intima/media ratio decreased significantly because of inhibition of SMC proliferation [56]. These data suggest that NO mediated by eNOS inhibits neointimal hyperplasia induced by vascular injury.
iNOS has been shown to be expressed in the SMCs after cuff-induced vascular injury in rabbits [57, 58], and iNOS-KO mice showed a significant reduction of neointimal thickening induced by cuff placement [59]. Unlike eNOS-deficient mice, iNOS-KO mice showed no reduction of the neointimal hyperplasia associated with mechanically induced endothelial denudation. Both the medial area and medial thickness were increased in the iNOS-KO mice after mechanically induced endothelial denudation [60]. Consistent with these data, Yogo et al. demonstrated that vascular remodeling, but not neointimal hyperplasia, after carotid artery ligation was increased in the iNOS-KO mice [55]. The differences in the procedure used to induce vascular injury may be related to the degree of neointimal hyperplasia and vascular remodeling in the iNOS-KO mice. Cuff placement evokes significant participation of inflammatory cells, including macrophages, in the adventitia as compared to the ligation model, as mentioned above. Increase of inflammation in the adventitia may be associated with increased iNOS expression levels, which may promote neointimal formation induced by cuff placement. Mechanically induced endothelial denudation and ligation may mainly induce migration of the SMCs from the media to the intima, resulting in increased neointimal hyperplasia.
Similar results were also observed in soluble epoxide hydrolase (sEH)/ApoE double-KO mice and inhibition of sEH by 12-(3-adamantan-1-yl-ureido)dodecanoic acid, which suppress metabolism of epoxyeicosatrienoic acids (EETs). sEH/ApoE double-KO mice or mice with inhibition of sEH showed significant reduction of the neointimal formation in the femoral artery cuff model, but not in the carotid artery ligation model. The expressions of proinflammatory genes were significantly reduced in the femoral arteries of the sEH/ApoE double-KO mice [61].
2.5.5 Roles of Inflammatory Cytokines in Cuff-Induced Neointimal Formation
Inflammation is considered as an important factor in human atherogenesis [8]. An inflammatory response to vascular injury, mediated by proinflammatory cytokines, influences the progression of neointimal formation and development of atherosclerotic lesions.
It has been demonstrated that the expressions of toll-like receptors (Tlrs) 2 and 4 are markedly enhanced in human atherosclerotic plaques and vascular adventitia [62]. Tlr4 serves as the receptor for bacterial lipopolysaccharides (LPS) and also recognizes cellular fibronectin, heat shock protein 60, and endogenous peptides that are produced in response to tissue injury [63]. In the adventitia, not all Tlr4-positive cells are colocalized with macrophages. Although application of LPS between the cuff and artery augmented the neointimal formation induced by cuff-induced injury in the WT mice, no such finding was observed in the Tlr4-KO mice [64]. Application of Pam3Cys-SK4, a synthetic Tlr2 ligand, significantly enhanced the neointimal formation induced by cuff placement in the femoral arteries of the WT mice. No such increase of the neointimal formation was observed in the Tlr2-KO mice. In ApoE-KO mice, application of Pam3Cys-SK4 led to a significant increase in the formation of atherosclerotic plaques [65]. Tlr2 stimulation produced significant induction of inflammatory cytokines in human adventitial fibroblasts in vitro. Treatment with cathepsin K (CatK), which is one of the most potent of mammalian collagenases, increased the mRNA levels of inflammatory cytokines, including Tlr2 and Tlr4. CatK-KO mice showed significantly reduced neointimal formation following cuff placement and ligation, accompanied by a decrease in the expression levels of Tlr2 and Tlr4 mRNA [66]. These findings provide evidence for a link between inflammatory cytokines in the adventitia and intimal lesion formation.
Although Tlr2/4 are expressed on the cell surface, Tlr7/9 are expressed on the endosomes. Tlr7/9 were detected at sites of post-interventional remodeling and accelerated atherosclerosis [67]. In hypercholesterolemic apolipoprotein E*3-Leiden mice, femoral artery cuff placement led to a strong increase of the Tlr7/9 expressions [68]. Blockade of Tlr7/9 with a dual antagonist reduced neointimal thickening and foam cell accumulation; the intima/media ratio was reduced by 64.5 % and luminal stenosis by 62.8 %. Application of the Tlr7/9 dual antagonist also reduced arterial wall inflammation, with reduced macrophage infiltration and altered serum IL-10 levels. Stimulation of cultured macrophages with Tlr7/9 ligands enhanced TNFα expression, which was decreased by coadministration of the Tlr7/9 antagonist. Furthermore, the antagonist abolished the Tlr7/9-enhanced LDL uptake. The antagonist also reduced oxidized LDL-induced foam cell formation, most likely not via decreased influx, but via increased efflux induced by increased IL-10 levels.
2.5.6 Role of RAS in Cuff-Induced Neointimal Formation
The renin-angiotensin-aldosterone system (RAAS) has various physiological actions such as vasoconstriction and is known to be involved in the development of hypertension and atherosclerosis [69, 70]. Specifically, modulation of the local RAAS may play a key role, just like that of the systemic RAS, in the development of cardiovascular diseases. Angiotensin (Ang) II is a peptide that exerts potent vasoconstrictive action via the AT1 receptor-MAPK pathway. Although human renin (hRN)/human angiotensinogen (hANG)-transgenic (Tg) mice showed increased blood pressure and medial thickening even in the absence of cuff placement, the hRN/hANG-Tg mice showed even more pronounced inflammatory vascular remodeling after cuff placement. Treatment with an AT1 blocker inhibited cuff-induced neointimal formation associated with reduced inflammation, but independently of the blood pressure change [71]. When the direct renin inhibitor aliskiren was administered to C57BL/6 mice via an osmotic pump, it inhibited cuff-induced vascular remodeling. The number of adherent leukocytes was increased in the cuff-injured mice not treated with aliskiren, where it was significantly reduced in the aliskiren-treated mice without any change of the blood pressure. Aliskiren decreased the adhesion of THP-1 cells to TNFα-stimulated human umbilical vein endothelial cells [72]. These data indicate that RAS activation augments neointimal hyperplasia induced by cuff placement via increased release of inflammatory cytokines.
Moreover, the ACE expression level increased in a time-dependent manner after cuff placement and was observed in the medial and neointimal layers and the adventitia of the cuffed arteries in FVB/N mice. The intima/media ratio after cuff placement was significantly decreased by ACE inhibitor treatment [73]. Ang II appears to be one of the factors exacerbating cuff-induced neointimal formation. In fact, AT1 receptor-KO mice showed decreased neointimal formation following cuff placement, accompanied by an increase of apoptotic cells among the SMCs [74]. Consistent with these data, the AT1-selective receptor blocker olmesartan suppressed cuff-induced neointimal formation via reducing ERK phosphorylation [75]. On the other hand, neointimal formation induced by cuff placement was increased in AT2 receptor-KO mice. The expressions of bcl-2 and bcl-xL mRNA, which are regulators of apoptosis, were enhanced in the AT2 receptor-KO mice showing enhanced neointimal formation [74].
Recently, in addition to the Ang II-AT1 receptor pathway, Ang-(1-7), which is synthesized from Ang I and Ang II mainly via ACE2 activity, has been reported to play a crucial role in vascular remodeling via Mas receptor activation [76, 77]. Mas-KO mice showed markedly increased neointimal formation after cuff placement, independently of the AT1 receptor. Treatment with Ang-(1-7) also suppressed neointimal formation, associated with suppression of vascular SMC proliferation, release of inflammatory cytokines and superoxide anion production in the injured artery. On the other hand, these inhibitory effects of Ang-(1-7) were less marked in the Mas-KO [78]. Interestingly, treatment with an AT1 receptor blocker inhibited neointimal formation induced by cuff placement, accompanied by a decrease in the expression levels of ACE2 and Mas mRNA and an increase in the expression of AT2 receptor mRNA. AT2 receptor-KO mice showed no reduction of the neointimal formation by treatment with Ang-(1-7). These results suggest that in addition to the activities of the ACE2/Ang-(1-7)/Mas axis, blockade of the AT1 receptor could enhance the activities of the ACE2/Ang-(1-7)/AT2 receptor axis and thereby inhibit neointimal formation induced by cuff placement.
2.5.7 Role of ROS in Cuff-Induced Neointimal Formation
Atherosclerosis is associated with increased production of reactive oxygen species (ROS) in the vessel [79]. The superoxide dismutases (SODs) are enzymes that catalyze the dismutation of superoxide anions to hydrogen peroxide; three isoenzymes of the SODs have been identified, namely, manganese SOD (MnSOD), which is localized in the mitochondria; copper/zinc SOD (Cu/ZnSOD), which is localized in the cytosol; and extracellular SOD (EC-SOD). The antioxidant enzyme Cu/ZnSOD metabolizes superoxide anions (O2-) in the vascular endothelial cells [80].
Although there was no difference in the degree of cuff-induced neointimal formation between the Cu/ZnSOD-KO and WT mice, the former showed a significant decrease in the intima/media ratio after cuff placement [81]. This increased medial SMCs in the Cu/ZnSOD-KO mice showed positive staining for SMemb/MHC-B, which is a useful molecular marker of embryonic-type SMCs. Moreover, the expression levels of TNFα, ICAM1, VCAM1, and iNOS in the media were higher in the Cu/ZnSOD-KO mice than in the WT mice, suggesting that Cu/ZnSOD-KO mice showed enhanced inflammation, expression of adhesion molecules, and altered structure of the media post-injury.
When an adenovirus vector expressing EC-SOD (AxCAEC-SOD) was injected between the cuff and the adventitia of the femoral arteries in rat models, neointimal formation was significantly reduced in the AxCAEC-SOD-transfected arteries [82]. Furthermore, proliferation of SMCs in the neointima and media was inhibited by EC-SOD treatment. Augmented iNOS expression, apoptosis, collagen content, and ROS generation in the vascular wall were also reduced by EC-SOD treatment. The amount of generation of ROS may have influence on the degree of medial thickening as well as neointimal formation induced by cuff placement.
2.5.8 Role of Insulin Signaling in Cuff-Induced Neointimal Formation
Insulin resistance has been reported to be associated with atherosclerosis; however, the underlying mechanism is still unknown [83]. Insulin resistance is caused by impaired insulin signaling induced by some factors. Insulin activates insulin receptor substrates (IRS) 1 and 2, which are expressed ubiquitously in various tissues, including the endothelial cells and SMCs, through the insulin receptor and the downstream signaling [84]. Both IRS1- and IRS2-KO mice show features of the metabolic syndrome, including insulin resistance, dyslipidemia, and hypertension. Although both genotypes of mice show a similar degree of insulin resistance, the IRS2-KO mice show more pronounced cuff-induced neointimal formation than the IRS1-KO mice, which in turn show more pronounced cuff-induced neointimal formation than the WT mice. IRS2 expression, but not IRS1 expression, is detected in the blood vessels [85]. Insulin resistance in the blood vessels is considered to exacerbate cuff-induced neointimal formation.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


