CHAPTER 8 Clinical Aspects of Seasonality in Mares
SEASONAL REPRODUCTIVE PATTERNS
Breeding (Ovulatory) Season
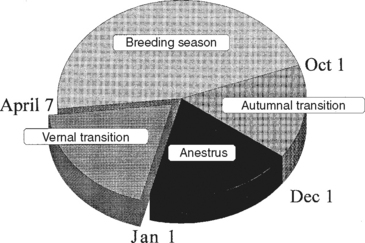
Equine breeding (or ovulatory) season is a time of reproductive competence occurring from mid spring through summer (Fig. 8-1). Nonpregnant mares experience numerous, repeated estrous cycles, ostensibly for the purpose of providing multiple successive opportunities to become pregnant. In other words, mares are seasonally polyestrous. Ovarian steroids (i.e., follicular estradiol and luteal progesterone) alternate dominance over the reproductive tract and sexual behavior in mares, setting suitable conditions for service, conception, and pregnancy.
Anestrus
Anestrus is a time of complete reproductive incompetence, marked by reduction in hypothalamic GnRH content1,2 and secretion3 with a consequent sharp reduction in LH and FSH secretion from the pituitary.2,4 Pituitary LH content is also dramatically reduced during anestrus.2,4 As a result of this hypothalamic-pituitary slowdown, ovaries undergo functional, if not morphologic, atrophy with only a few follicles over 5 to 10 mm observed. Lack of follicular development is echoed by virtual loss of ovarian steroids in peripheral circulation, and progesterone and estradiol become essentially undetectable.6–8 During anestrus, mares’ sexual behavior toward stallions becomes more passive. They may not object to a stallion’s presence and may even permit mounting, but intromission is often prevented by holding their tail down.9
Vernal Transition
Springtime (or vernal) transition from anestrus into the ovulatory season is economically and scientifically important. Desire, throughout much of the equine industry, for early-born foals conflicts with innate physiologic regulation of reproductive competence. Furthermore, vernal transition is filled with ambiguous signs that disguise the underlying process, confuse breeders, and lead to low reproductive efficiency. Owing to a lack of understanding, breeders often cover mares based solely on behavioral signs of estrus and without regard for their state of reproductive competence. On the other hand, vernal transition has been shown to follow a well-regimented series of steps leading to resumption of reproductive activity (Fig. 8-2). Vernal transition is the most precisely regulated of the annual reproductive rhythms, with relatively little variation among mares in timing of their first ovulation of the year. One of the first events in vernal transition is resumption, albeit at a reduced level compared with the ovulatory season, of GnRH secretion by the hypothalamus. Examination of hypothalamic GnRH content1,2,10 and secretion3 indicates the GnRH neural network increases its activity shortly after winter solstice. This increased GnRH secretion appears to be maintained throughout much of vernal transition. In response to increased GnRH, FSH secretion resumes. Though speculative, apparently the gene-regulating FSH subunit synthesis remains functional during anestrus and the lack of FSH secretion reflects a lack of GnRH secretion rather than pituitary FSH content. Nevertheless, an increase in FSH secretion is temporally associated with increased GnRH.

Ovaries, quiescent until this point, respond by developing new follicles. Follicular growth pattern during vernal transition is relatively predictable, with increased size and numbers of follicles. Of major importance, the first several follicles that develop during vernal transition generally do not ovulate, although they may reach normal preovulatory size (>30 mm). This is one of the major events contributing to reproductive inefficiency during vernal transition, as it is difficult to know whether a given follicle is competent and will ovulate. After several (3.7 ± 0.9) of these transition follicles develop and regress without ovulating, a competent follicle develops.8 Although it is not yet known what factors contribute to eventual development of the first competent follicle, it is clear that this follicle, destined to be the first ovulatory follicle of the year, is reasonably steroidogenically competent. Development of the first ovulatory follicle is accompanied by a surge of estradiol in peripheral plasma followed a few days later by a surge in LH. The first ovulation of the year follows, ending vernal transition.
CLINICAL ASPECTS OF SEASONALITY
Challenges Seasonality Presents to Breeders and Veterinarians
A circannual rhythm in mares entrained by environmental cues ensures foaling during spring and summer. However, many breed associations observe January 1 as the official birth date of foals (in order to be eligible for registration in their respective association) born in the same calendar year. Therefore, seasonality in mares presents breeders and veterinarians with a major challenge to produce foals early. Though other environmental factors may also exert some influence on the annual reproductive rhythm in mares,11–20 photoperiod is widely accepted as the driving force.21 Increased photoperiod has long been recognized as a means of returning seasonally anestrous mares to reproductive competence sooner than would occur with natural day length.22 However, increasing day length with artificial light does not shorten transition but merely causes it to begin earlier in the year.23 Whether transition is brought about by natural or artificial light, it is a lengthy process lasting 6 to 12 weeks or more and there is considerable variation in interval from start of treatment to first ovulation among mares.24,25 Horse breeders would benefit from a treatment or management protocol that would not only shift the beginning of the ovulatory season to a point earlier in the year but would also shorten the length of vernal transition.
Though mares in vernal transition proceed through a regimented series of steps leading to resumption of reproductive activity (Fig. 8-2), ambiguous signs characteristic of the phase are difficult to interpret and present considerable challenges for breeders and veterinarians managing transitional mares. Unknowledgeable breeders often equate behavioral signs of estrus with the ovulatory season and cover mares repeatedly during irregular and often lengthy periods of sexual receptivity. Even when follicular growth and development are monitored with behavior, difficulty in predicting the first ovulation of the year can further contribute to low reproductive efficiency.
Photostimulation of Reproductive Resurgence
In 1947, Burkhardt was the first to report early breeding of mares after doubling their daylength by adding artificial light.22 Since then, a variety of photoexposure regimens have been used in mares to stimulate an early return to reproductive competence.26–28 Traditionally, horse breeders exposed mares to a 16-hour fixed-length photoperiod throughout transition using a combination of natural and artificial light. Although very effective, such a long photoperiod can involve considerable expense and inconvenience. Other options available today may help breeders reduce overall expense and inconvenience of their photostimulation program. Extending natural day length with as little as 2 to 3 hours of additional artificial light was sufficient to accelerate onset of the ovulatory season.21,24 Of interest, adding light prior to sunrise was completely ineffective, whereas adding light at sunset, thus extending the day, was very effective.21,24 Guidelines for providing artificial light in a box stall or paddock setting have been reported.21,29 Another effective method of providing photostimulation is the so-called “night-interruption” or “pulse lighting” approach.30 Artificial light is applied for 1 to 2 hours during a photosensitive phase 9.5 hours after the beginning of darkness. The disadvantage of this approach is that “pulse” timing must be adjusted to changing onset of darkness or onset of darkness must be controlled artificially.25
Timing the start of a photostimulation program is important to effectively shift transition. Ideally mares enter the ovulatory season 1 or more cycles before intended breeding, thus eliminating the need to predict the first of ovulation and allowing ample opportunity to evaluate and prepare, if necessary, their uteri for conception and pregnancy. To effectively gain an advantage over natural photoperiod, artificial light should begin no later than winter solstice.30 However, an even greater advantage may be possible when artificial light is applied as soon as 5 to 7 weeks before winter solstice.24,29 One author’s (GJN) clinical experience supports that assertion. Breeders waiting until January to expose their mares to increased day length will not realize any advantage over exposing them to natural photoperiod.24,31 Traditionally it was recommended that artificial lights be continued until late spring,28 presumably until natural day length was sufficiently long to prevent an abrupt differential in photostimulation that potentially could interfere with cyclic activity. However, it was recently reported that 14.5 hours of light applied for only 35 days beginning at winter solstice was sufficient to advance the ovulatory season.28 The mares reached their first ovulation in a similar time as other mares exposed to an extended daily photoperiod throughout the winter and spring. Further, they remained reproductively active after artificial light was discontinued even though natural day length at the time was considerably shorter than the artificial photoperiod.
Dating back to the first report of photostimulation in mares, traditionally a 100 W incandescent light bulb, providing luminal intensity of 10 to 12 foot-candles (approximately 100 lux), has been recommended for use in a 12 × 12 box stall to stimulate onset of reproductive activity.22,29 However, studies have investigated light intensities down to 10 lux (approximately 1 foot-candle).21 One report indicated that light intensity of only 10 lux, when applied daily, for 14.5 hours, throughout the winter and spring, was sufficient to advance the onset of reproductive activity.28 Little work has been done to determine the optimal spectral wavelength of light used for photostimulation in mares. Nevertheless, recognizing the eye as the primary photo receptor and maximal photochemical sensitivity being in the midrange (approximately 550 nm), it was suggested that light sources with emission ranges that include the midrange will work.21
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


