CHAPTER 63 Cestodes
ETIOLOGY
Three species within the family Anoplocephalidae (Kholodkovskii, 1902) infect the gastrointestinal (GI) tract of horses and donkeys: Anoplocephala perfoliata (Goeze, 1782; Blanchard, 1848), Anoplocephala magna (Abildgaard, 1789; Sprengel, 1905), and Anoplocephaloides mamillana (Mehlis, 1831; Rausch, 1976; Schmidt, 1986). All members of the family Anoplocephalidae are similar in that the scolex is devoid of rostellum, hooks, or hooklets during all stages of development, and the suckers are unarmed.1,2 Within this family, four subfamilies have been described based on type of uterine development.2 All members of the subfamily Anoplocephalinae (Blanchard, 1891), including the three equine tapeworm species, can be defined by the following characteristics: morphologically, the uterus persists in gravid proglottids, and biologically, members have a cysticercoid larval type that occurs within the hemocele of oribatid mites.2,3
Morphology
Distinguishing characteristics of adult Anoplocephala perfoliata (Fig. 63-1, A) include the following: (1) the length of gravid adult specimens is usually 25 to 40 mm but may reach 80 mm; (2) the width of the body is generally between 8 and 14 mm; and (3) the scolex or holdfast organ is distinct and much smaller than the body, measuring only 2 to 3 mm.1
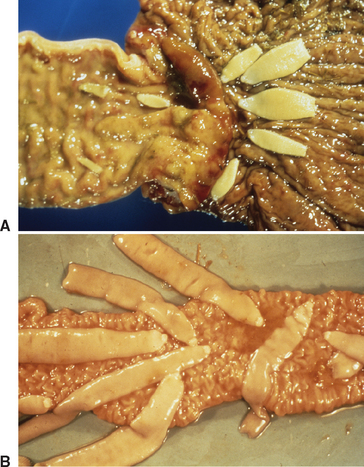
The scolex of A. perfoliata has four ear-shaped lappets measuring 0.5 to 1.0 mm that are situated posterior to the four apical muscular suckers.4 In contrast, Anoplocephala magna is generally larger, measuring up to 80 cm in length and 2.5 cm in width, with a scolex 4 to 6 mm wide (Fig. 63-1, B), and Anoplocephaloides mamillana is smaller, measuring 6 to 50 mm in length and 4 to 6 mm in width.3 The scolices of the latter two species do not have lappets as does A. perfoliata.
The morphology of proglottids of A. perfoliata has been described extensively.1 Individual proglottids are always much wider than long. Each proglottid is hermaphroditic, containing a single set of both male and female reproductive organs. Each proglottid has a single genital apparatus; the apertures are unilateral and are found in the cranial half of the lateral margin. The gravid uterus is transverse, large, saclike, and lobed.3 The female reproductive system also consists of a poral ovary, a vagina posterior to the cirrus sac, and a seminal receptacle. Both internal and external seminal vesicles are present as part of the male reproductive system, and the testes are scattered throughout the medulla.2 Each proglottid also contains a muscular system, a tegument, and an excretory system.
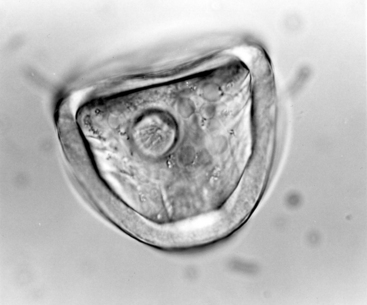
The morphology of mature eggs of A. perfoliata is unique (Fig. 63-2). The eggs are 65 to 80 μm in diameter, whereas eggs of A. magna measure 50 to 60 μm in diameter and Anoplocephaloides mamillana about 51 by 37 μm.3 A. perfoliata eggs are round to D shaped, with an outer vitelline membrane and a thick (8-10 μm), dark, albuminous middle shell. The innermost membrane is flame or pear shaped and consists of a chitinous pyriform apparatus. The length of the pyriform apparatus is approximately equal to the radius of the egg, measuring about 48 μm. This pyriform apparatus in turn contains the hexacanth embryo characteristic of cyclophyllidean cestode eggs.1,5 The diameter of the embryo measures approximately 16 μm. Eggs seen on fecal flotation often have an amber cast resulting from contact with excreta; however, eggs dissected from gravid proglottids of the adult tapeworm are colorless.1
The morphology of anoplocephalid larval development is very similar among many species. In fact, it is difficult to impossible to determine the genus from studying the cysticercoid.1 The morphology of the larval stages of Anoplocephala perfoliata within the intermediate host has been described.6 However, literature generally presents a detailed ontogenesis of the tapeworm of sheep and goats, Moniezia expansa rather than this parasite.1,7 Briefly, the stages of ontogenesis of A. perfoliata begin after the tapeworm egg has been ingested. The oncosphere emerges and penetrates the intestinal wall of the oribatid mite. The oncosphere appears in the mite’s body cavity within 48 hours and is very motile and active for several days to weeks. The second stage of development is the “large sphere” stage, in which the oncosphere form is lost and the larva becomes immobile. During this period, the oncosphere increases in size and undergoes internal reorganization of the organs. The six hooks lose positioning and move to the tail of the larva while the body of the larva fills with round cells. The third stage of growth within the mite’s hemocoele is the “extended larva” stage, in which the body lengthens. The fourth stage is called the “segmented larva” stage. In this phase of development, the body continues to elongate but also divides into two parts that are separated by a constriction. The portion anterior to the constriction has four suckers, which will become the scolex of the tapeworm, and the portion posterior to the constriction is a spherical capsule. Embryonic hooks are positioned caudal to the capsule. During the fifth stage of larval development, the anterior portion of the larva invaginates into the posterior capsule portion. The “cysticercoid stage” is the final stage of larval development. Morphologically, the body is spherical with a dense cuticle. Within the cuticle is the scolex, with four suckers. The wall of the cyst is stratified. At this phase of development, the larva is infective for the final host. In six experimentally infected Scheloribates spp. the average size of the A. perfoliata cysticercoids was 141 by 119 μm.8
Life Cycle
The life cycle of the three tapeworm species that infect the GI tract of horses and donkey are indirect because they require an intermediate host as well as a definitive host. Stunkard9 completed the developmental cycle of Anoplocephaline cestodes in 1937 when he discovered that certain members of one genus of oribatid mites (Acari: Oribatida), Galumna spp., could serve as the intermediate hosts of the sheep tapeworm, Moniezia expansa, a closely related cyclophyllidean that infects ruminants. A few years later, Bashkirova6 determined the complete developmental cycle of Anoplocephala perfoliata. With an indirect life cycle, the prevalence of cestode infection in the intermediate host is low, whereas it is high in the definitive host.10 Intermediate host specificity is low in the various anoplocephaline cestode species, and many species of oribatid mites may become infected.11
Adult A. perfoliata parasites attach near the ileocecal valve of horses, whereas A. magna and Anoplocephaloides mamillana attach in the small intestines of the horse’s digestive tract. Horses with patent infections shed tapeworm eggs in their feces. The proglottids of A. perfoliata are broken up by digestion during transit through the large intestines, and thus only eggs are passed in the feces.12,13 After a gravid proglottid is shed, more than 48 hours may elapse before its ova are passed in the feces.13
Survival time of an infective egg on pasture is important because it potentially allows for an improved chance of exposure to the intermediate host. Little is known about the longevity of A. perfoliata eggs in the environment. It has been hypothesized that cestode egg survival may be shortened in tropical climates.14 However, the infectivity of cestode eggs over time in natural conditions has not been determined. Stunkard15 believed that the redistribution of the anoplocephaline tapeworm eggs by rain into the upper layers of the soil would allow the eggs to remain viable for a longer period and enable the intermediate hosts in the soil to encounter the eggs, pick them up, and feed on them.
The life cycle continues when oribatid mites ingest viable eggs of A. perfoliata on the pasture. Oribatid mites are free-living mites found on herbage and in the soil of pastures. Because the cestode eggs are presumably too large to be accidentally eaten, the intermediate host may interpret them as prospective or preferred food.16 Mackiewicz10 hypothesized that Anoplocephaline tapeworm eggs may use chemoattraction to increase the likelihood of being eaten by the oribatid mites. This would especially hold true if the tapeworm eggs were eaten as a food item rather than as a food contaminant. On the other hand, the oribatid intermediate hosts and A. perfoliata eggs may be so abundant that chance encounters between the egg and mite may be the sole strategy of transmission.10,11
Whatever the transmission strategy, once ingested by the intermediate host, the larva or oncosphere is freed from within the tapeworm eggshell or embryophore, presumably through digestion. Activation factors that stimulate the oncosphere to tear through the intestinal wall using its hooks are unknown.17 Once in the mite’s body cavity, the development time to an infective cysticercoid within the invertebrate intermediate host is variable. Growth of the cysticercoid within the mite’s hemocele depends on environmental conditions, especially temperature. The infective cysticercoid stage is formed within 8 to 20 weeks under natural conditions. Once the cysticercoid is fully developed and infective within the oribatid mite, it is ingested by a grazing horse.
The infective cysticercoid has a scolex with four fully developed suckers. It is assumed that the excysted larvae of A. perfoliata move along the GI tract with ingesta until reaching the ileocecal valve area, at which point the larvae attach to the GI mucosa. Behind the scolex of the larvae, germinal cells will multiply through proglottid development or asexual reproduction to produce the proglottids of the adult tapeworm.15,18,19 The caudal end of the larvae contains the excretory pore that becomes the terminal segment of the adult tapeworm.15 The prepatent period following ingestion of an infected oribatid mite is 6 to 16 weeks.5,20,21,22
No actual data are available on the life span of adult A. perfoliata, but the life span of an adult cestode may vary from a few months to several years.18,23 A basic cestode life cycle strategy, based on repeated production of egg-laden proglottids, infers a long adult cyclophyllidean life span that often lasts for years or as long as the definitive host lives.10,18,24 Furthermore, there is selection for repeated production of proglottids and eggs and high fecundity when the pre-reproductive life span is long.18 In the case of A. perfoliata, this pre-reproductive life span may be 1 to 1½ years while the cysticercoid is in the oribatid mite.25,26
EPIDEMIOLOGY
The distribution and prevalence of A. perfoliata are high enough to cause concern among both horse owners and veterinarians. A. perfoliata is found worldwide and is currently accepted as the most common and the most pathogenic of the equine tapeworm species.3,22,27–31
The prevalence of A. perfoliata in North America has been reported extensively. Many epidemiologic studies report prevalence of infection at necropsy. In 1979, necropsy data collected from eight states, including Kentucky, found that 18% of foals and 26% of adult horses were infected with tapeworms.32 The most common tapeworm identified was Anoplocephala magna. Other studies reported prevalences of A. perfoliata in the United States that vary from 13% to 54% and worldwide from 14% to 81.5%* (Box 63-1). Why different regions have such varying prevalence rates for these parasites is not known. The differing rates could be caused by differences in pasture type (thus creating a better or worse environment for the intermediate hosts), pasture stocking rates, climate, or other management or environmental factors. Even with the variation in prevalence, there seems to be a trend toward higher prevalence of infection in countries with temperate climates.13
Box 63-1 Prevalence (%) of Anoplocephala perfoliata in United States (US) and Other Countries
| North Carolina (US) | 13% |
| Kentucky (US) | 52%-54% |
| Louisiana (US) | 47% |
| New England (US) | 53% |
| Ohio (US) | 18% |
| Canada | 14% |
| New Zealand | 81.5% |
| England | 31%-69% |
| The Netherlands | 21% |
| Sweden | 65% |
Many parasitologists have speculated about the factors that may have caused an increase in reports of A. perfoliata infection. Edwards44 and Geering and Johnson45 proposed that extensive use of ivermectin, which was new to the market, removed the nematode parasites with greater efficacy than drugs previously available. This selective removal of other intestinal parasites allowed tapeworms to flourish because of lack of competition, thus the increase in prevalence of A. perfoliata in recent decades. This hypothesis was refuted by French et al.,46
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


