Fig. 2.1
The location of pig’s liver and gallbladder
Four lobes are surrounded by a capsule of fibrous connective tissue called Glisson’s capsule. The Glisson’s capsule in turn is covered by the visceral peritoneum (tunica serosa), except where the liver adheres directly to the abdominal wall or other organs. The parenchyma of the liver is divided into lobules, which are incompletely partitioned by septa from Glisson’s capsule (Ekataksin and Wake 1991). The parenchyma within the lobules is supported only by fine reticular fibers (which are discernible only with special preparations).
The blood vessels supplying the liver (portal vein and hepatic artery) enter at the hilum (or porta hepatis), from which the common bile duct (carrying bile secreted by the liver) and lymphatic vessels also leave (Fig. 2.2).
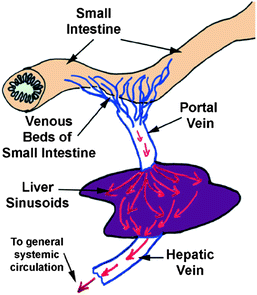
Fig. 2.2
The liver’s structure and anatomical placement
Within the liver sinusoids, the oxygen-poor but nutrient rich blood from the portal vein mixes with the oxygenated blood from the hepatic artery. From the sinusoids, the blood enters a system of veins which converge to form the hepatic veins. The hepatic veins follow a course independent of the portal vessels and enter the inferior vena cava. Liver cells, or hepatocytes, are large polygonal cells, usually tetraploid and often binucleate in the adult (Sen and Jalan 2005). Each nucleus has two or more nucleoli. The average life span of liver cells is 5 months. They contain abundant rough endoplasmic reticulum and mitochondria, large deposits of glycogen and lipid droplets of various sizes, and several small elaborate Golgi complexes. They also contain many peroxisomes, a variable amount of smooth endoplasmic reticulum and lysosomes (Vega 2004). In standard histological preparations, liver cells usually appear vacuolated because the glycogen and lipids are removed during processing. Liver cells are capable of considerable regeneration when liver substance is lost (Harada et al. 2009).
2.3.2 The Function of the Liver
The liver is extremely versatile, having both endocrine and exocrine functions. Liver is involved in numerous metabolic activities and acts as a storage depot. The list below is an attempt to categorize its numerous and diverse activities.
Blood Supply
As mentioned above, the liver receives a dual blood supply. The hepatic portal vein provides the majority of the incoming blood, the rest coming from the hepatic artery. The blood from the portal vein has already supplied the small intestine, pancreas, and spleen, and is largely deoxygenated. It contains nutrients and noxious materials absorbed in the intestine. The blood from the hepatic artery supplies the liver with oxygen. Because the blood from the two sources intermingles as it perfuses the liver cells, these cells must carry out their many activities under low oxygen conditions that most cells would not be able to tolerate.
The Protein and Lipid Synthesis
The liver synthesizes a number of proteins, as suggested by its abundant rER and Golgi. One of the most important of these is albumins (responsible for several functions including colloid osmotic pressure), the protein parts of several kinds of lipoproteins, nonimmune alpha and beta globulins, prothrombin, and numerous glycoproteins, including fibronectin. These proteins are considered as endocrine secretions because they are released directly from the liver cells into the blood supply. The liver also synthesizes cholesterol and the lipid portion of lipoproteins. The enzymes for these activities are found in the sER.
Metabolic Functions
The liver is involved in a wide range of metabolic activities. Among its major metabolic functions are gluconeogenesis (the formation of glucose from non-carbohydrate precursors), and the deamination of amino acids. In addition to the proteins listed above, the liver also releases a number of other products directly into the blood as endocrine secretions. These include products of carbohydrate metabolism, e.g., glucose released from stored glycogen, and modified secretions from other organs, e.g., triiodothyronine (T3).
Storage of Metabolites
Liver cells store large deposits of glucose in the form of glycogen, which can be stained by the periodic acid-Schiff or PAS procedure. They also store lipids in the form of droplets of varying sizes. The latter can be revealed with Sudan staining after tissue fixation. The removal of these materials in routine preparations gives liver cells a foamy or vacuolated appearance. Lysosomes within hepatocytes may be a normal storage site for iron, in a form of a ferritin complex. Excessive amounts of iron may accumulate in cells as an unusable yellowish-brown pigment called hemosiderin, a partly denatured form of ferritin. Liver lysosomes also contain pigment granules (lipofuscin) and partially digested cytoplasmic organelles. The liver also stores vitamins, especially vitamin A, which is transported from the liver to the retina to be used in the formation of visual pigments. Most of the vitamin A is not stored in the hepatocytes, however, but in special fat-storing cells, called Ito cells, within the sinusoids. Ito cells also store the other three fat-soluble vitamins, D, E, and K, as well as vitamin B12.
Detoxification and Inactivation of Noxious Substances
The liver is, after the gastrointestinal tract, the first organ to receive metabolic substrates and nutrients from the intestine, but it is also the first to receive any toxic substances, for instance carcinogens or drugs that have been ingested. The sER of liver cells contains enzymes involved in the degradation and conjugation of toxins (in addition to those responsible for synthesizing cholesterol and the lipid portion of lipoproteins). Under conditions of challenge by drugs, toxins or metabolic stimulants, the sER becomes the predominant organelle. Stimulation by one drug, e.g., alcohol, enhances its ability to detoxify some other compounds. On the other hand, some toxins can get metabolized by the liver to even more damaging compounds.
2.4 The Structure and Function of the Gallbladder in Pigs
The gallbladder is one of the most dispensable organs in mammal. The gallbladder is a small sac and is the storage depot for bile. Bile is only concentrated in the gallbladder. The gallbladder is located at the back part surface of the liver (Fig. 2.1). It is connected with the liver via the cystic duct and the hepatic bile duct. Together they form the common bile duct, which leads into the small intestine.
2.4.1 The Structure of the Gallbladder in Pigs
The gallbladder is a tubular organ. Its tunica mucosa is a very regular simple columnar epithelium, with no goblet cells or glands. The deep folds in the bottom of the mucosa, which are often cut in cross section, sometimes look like “glands,” but they have no secretory activity. There is a scanty lamina propria, and a thin tunica muscularis. There is no muscularis mucosae, and this is one of the “landmarks” for this organ. The gallbladder is only partially covered by a fold of peritoneum; its opposite side is seated in a pocket in the surface of the liver. Bile ducts carry bile to the gall bladder for temporary storage.
The low magnification image above shows the walls of the gallbladder (Fig. 2.3). They vary in height depending on how full the bladder is. At higher magnification, the nature of the gallbladder epithelium is revealed: a simple columnar form, containing no goblet cells. Though these cells do have a few microvilli on their free surfaces, they cannot be considered as real brush borders. There is no muscularis mucosae in these structures, and the tunica muscularis is stringy and scanty with collagen fibers. Since it has only the function of squirting bile into the duct that carries it to the duodenum, it appears little developed.
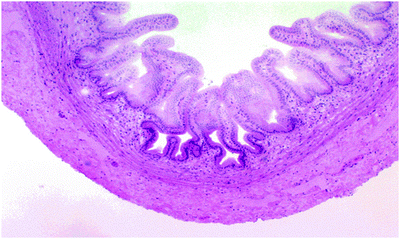
Fig. 2.3
The wall of the gallbladder at the low magnification image
2.4.2 The Function of the Gallbladder in Pigs
Bile is continuously formed in bile canaliculi in the liver hepatocytes, were it is collected and secreted into the hepatic bile duct. Bile is a complex fluid containing water, electrolytes, and a battery of organic molecules including bile acids (water-soluble derivatives of cholesterol), cholesterol, phospholipids, and bilirubin (Mcmaster and Elman 1926). The rate of bile fluid secretion is dependent on the circadian rhythm of the animal.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


