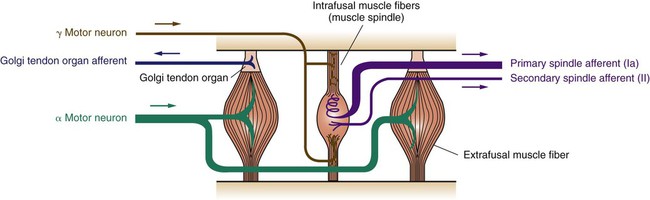
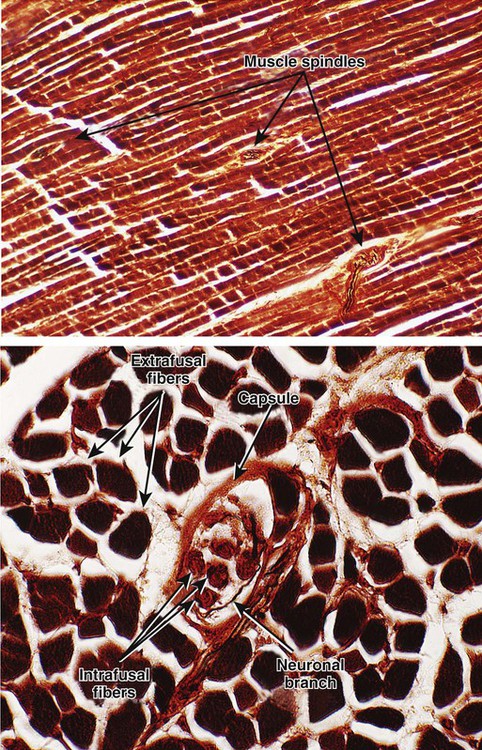
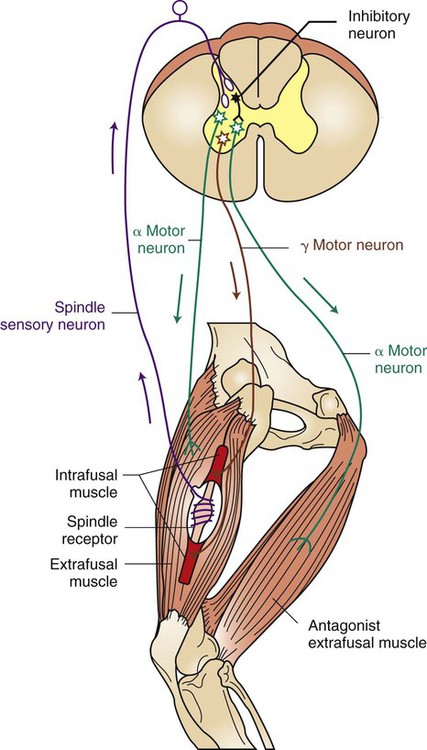
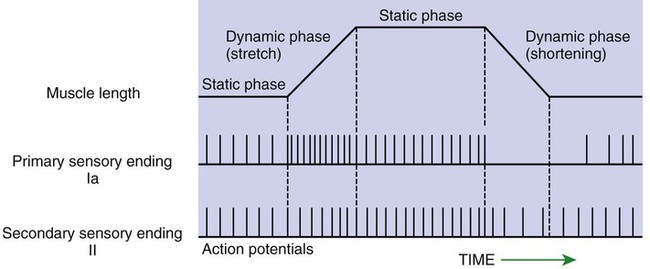
1. The muscle spindle stretch receptor is an encapsulated organ of specialized muscle fibers with separate motor and sensory innervations. 2. The muscle spindle conveys information about muscle length to the central nervous system. 3. Muscle stretch and action potentials along spindle sensory neurons lead to reflex contraction of the extrafusal muscle fibers. 4. The central nervous system can control spindle sensitivity directly through the gamma (γ) motor neurons. 5. The Golgi tendon organ lies in series between muscle and tendon and detects muscle tension. 6. Free (non-organ) sensory receptors in joints and muscles can provide information about joint position, joint movement, and pain-inducing stimuli of joints and muscles. Movement, characteristic of all animals, is the end product of skeletal muscle contraction. It is orchestrated by the central nervous system (CNS) through its control of the motor unit (see Chapter 6). To control body movement appropriately, the CNS must (1) assess the effect of gravity on the many muscles of the body, (2) determine the initial position of the body parts to be moved, and (3) detect any discrepancy between the intended movement and the movement that actually occurs. When such discrepancies are detected, appropriate adjustments can be made. Two important receptor systems have evolved in the skeletal muscles of mammals to provide the CNS with the aforementioned information: the muscle spindle and the Golgi tendon organ (Figure 8-1). The muscle spindles, arranged in parallel to the contracting skeletal muscle fibers, provide information about muscle length. The Golgi tendon organ, arranged in series with the contracting skeletal muscle fibers, detects muscle tension. This chapter discusses the anatomy and physiology of these two receptor organs; Chapter 10 discusses how the CNS uses the information gathered from these organs to coordinate posture and locomotion. Some of this information is used in reflex arcs of the type described in Chapter 7. The muscle spindle is an encapsulated group of about 3 to 12 small, slender, specialized skeletal muscle fibers (Figure 8-2). Because their capsule is spindle shaped, or fusiform, these muscle fibers are called intrafusal muscle fibers. The muscle fibers that cause physical shortening of the muscle (the majority of muscle fibers in a muscle belly), located outside the capsule, are called extrafusal muscle fibers. Extrafusal muscle fibers often span the length of the gross muscle from origin to insertion tendon; intrafusal muscle fibers and their capsules are much shorter (about 4 to 10 mm long). In addition, the intrafusal muscle fiber endings are attached to the extracellular matrix of, and lie in parallel to, the extrafusal muscle fibers. Therefore, if the muscle is stretched, lengthening the extrafusal muscle fibers, the intrafusal fibers of the muscle spindle are also stretched. Spindle sensory neurons arise from the equatorial region of the intrafusal muscle fibers and carry action potentials from the spindle to the CNS by way of the peripheral nerves. These CNS afferents enter the spinal cord through the dorsal roots (Figure 8-3). The contractile, polar regions of the intrafusal muscle fibers are innervated by motor neurons called gamma (γ) motor neurons. Extrafusal muscle fibers—the muscle fibers that cause the physical shortening of the muscle—are supplied by a different population of motor neurons (those that comprise the motor units) called alpha (α) motor neurons. Although γ motor neurons go to intrafusal muscle fibers and α motor neurons go to the extrafusal muscle fibers, these CNS efferents both have their cell bodies in the ventral horn of the spinal cord, and their axons leave through the ventral roots. As noted, stretching a muscle can stretch the intrafusal fibers of the muscle spindle that lie parallel to the extrafusal muscle fibers. Stretching (lengthening) the equatorial segment of the intrafusal muscle fiber generates action potentials along the spindle sensory neurons. As the equatorial segment is lengthened, it is believed that stretch-sensitive ion channels open on the sensory neurons, leading to membrane depolarization and action potential generation. Action potentials are generated along the sensory neuron output of the muscle spindle in proportion to the amount of lengthening of the middle of the intrafusal muscle fibers. There are actually subclasses of intrafusal fibers (dynamic nuclear bag, static nuclear bag, and nuclear chain) and spindle sensory neurons (Ia, II). This variety allows the sensory neuron output of the spindle to detect not only a change in length during the dynamic phase of muscle lengthening, but also the rate of lengthening, as well as the steady-state length of the muscle as the animal holds the joint still (Figure 8-4). When a muscle is held at a constant length, such as when a joint is held still, there is usually enough stretch of the muscle’s spindles to produce a steady frequency of action potential discharge in the sensory neuron output of the muscle spindle. This steady-state discharge gives the muscle spindle organ the ability to inform the CNS not only about a subsequent lengthening of the muscle, which would produce a proportional increase in action potential discharge, but about a shortening of the muscle as well, which would produce a proportional decrease in action potential discharge from the steady state (see Figure 8-4).
Skeletal Muscle Receptor Organs
The Muscle Spindle Stretch Receptor Is an Encapsulated Organ of Specialized Muscle Fibers with Separate Motor and Sensory Innervations
The Muscle Spindle Conveys Information About Muscle Length to the Central Nervous System
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


